
This species is highly variable in morphology, which can manifest by season as well as habitat. In its winter to early summer morphology some NW Atlantic specimens develop a corymbose morphology (Image A), while late summer into early autumn plants, as well as specimens from some habitats (e.g. deep subtidal off PE), typically lack this appearance (Image B). The former and latter recall the morphology of Rhodomela confervoides (Hudson) P.C.Silva (Image C) and Rhodomela lycopodioides (Image D) in NE Atlantic waters, respectively. Plants typically range from 6 to 36 cm in height with several erect axes arising from a basal disc with older axes ~1 mm in diameter (Images A-B). Branching is typically spiralled being sparser towards the base, with branches of successive orders becoming shorter and thinner (Image A). Thalli are uniaxial with an obvious, rounded apical cell (Image E). Although construction is polysiphonous, this is obscured by a cortical covering in whole mount (Image F). With some luck, a cross section will reveal the central rosette pattern of this species, which consists of a central axial filament, 40-60 µm in diam., surrounded by 6(-7) periaxial cells of larger dimensions, 90-140 µm long, these further surrounded by inner cortical cells of similar dimensions that gradually reduce in size to the outer cortex (Image G; mature thallus region). Long section also reveals the similarity in appearance and length (perpendicular to the main axis) of the periaxial cells (typically too tall to measure accurately) and inner cortical cells, the latter as much as 900 µm tall (parallel to the main axis) (Image H; mature thallus region). The similarity in size and shape of the periaxial and inner cortical cells in cross section, combined with a lack of certainty as to which cell is the axial cell in some sections, can make it difficult to identify the true central rosette. In Image I what appears to be axial cell and associated rosette is not; the axial cell being smaller and offset. Another section from this same specimen clearly reveals the smaller central axial cell surrounded by six periaxial cells (Image J). Near the tips of female thalli numerous procarps (female reproductive unit) develop low on fertile trichoblasts and are housed in young pericarps (Image K) from which the receptive trichogyne extends (Image L). Mature cystocarps are lateral, slightly urceolate to spherical, 420-440 µm wide by 475-490 µm tall, with an apical ostiole (Image M). Tetrasporangia are tetrahedrally divided, typically in pairs, but one partner can develop in advance of the other, spherical to oval, 76-88 µm wide by 98-102 µm tall (Image N).
Genetically verified records extend from RI to Hudson Bay in the Canadian Arctic. Mid intertidal pools to subtidal (20 m) on various hard substrata, this species accounts for all previous records of Rhodomela confervoides Hudson P.C.Silva (e.g. Image A) and all southerly (typically below NL) records of Rhodomela lycopodioides (e.g. Image B) in our flora. This species is genetically distinct from true Rhodomela confervoides Hudson P.C.Silva in Europe (Image C) and is closely allied with European Rhodomela lycopodioides (Image D) with which it is likely conspecific – taxonomic work is ongoing. There are no genetic matches for true Rhodomela confervoides Hudson P.C.Silva in our flora. Our species overlaps in range with Rhodomela virgata Kjellman and Rhodomela tenuissima (Ruprecht) Kjellman, but these both have a marked differentiation between the periaxial cells and the inner cortical cells such that the central rosette is easily observed in most sections (mature thallus regions). Additionally, Rhodomela tenuissima (Ruprecht) Kjellman has a large central axial cell, 75-100 µm, compared to these other two species.
 Image A. Corymbose tips of the “Rhodomela confervoides Hudson P.C.Silva” morphology from Welch’s Cove, NB (GWS043281).
Image A. Corymbose tips of the “Rhodomela confervoides Hudson P.C.Silva” morphology from Welch’s Cove, NB (GWS043281).
 Image B. More typically “Rhodomela lycopodioides” morphology from the deep subtidal (15 m) off Naufrage, PE (GWS041511).
Image B. More typically “Rhodomela lycopodioides” morphology from the deep subtidal (15 m) off Naufrage, PE (GWS041511).
 Image C. Bona fide Rhodomela confervoides Hudson P.C.Silva from low intertidal pools on rock near Bergen, Norway (2016_BIO309A_58).
Image C. Bona fide Rhodomela confervoides Hudson P.C.Silva from low intertidal pools on rock near Bergen, Norway (2016_BIO309A_58).
 Image D. Bona fide Rhodomela lycopodioides from a dredge (5-10 m) near Bergen, Norway (GWS040946).
Image D. Bona fide Rhodomela lycopodioides from a dredge (5-10 m) near Bergen, Norway (GWS040946).
 Image E. Rounded apical cell at branch tip (low intertidal pool; Musquash Head Light, NB; GWS044253).
Image E. Rounded apical cell at branch tip (low intertidal pool; Musquash Head Light, NB; GWS044253).
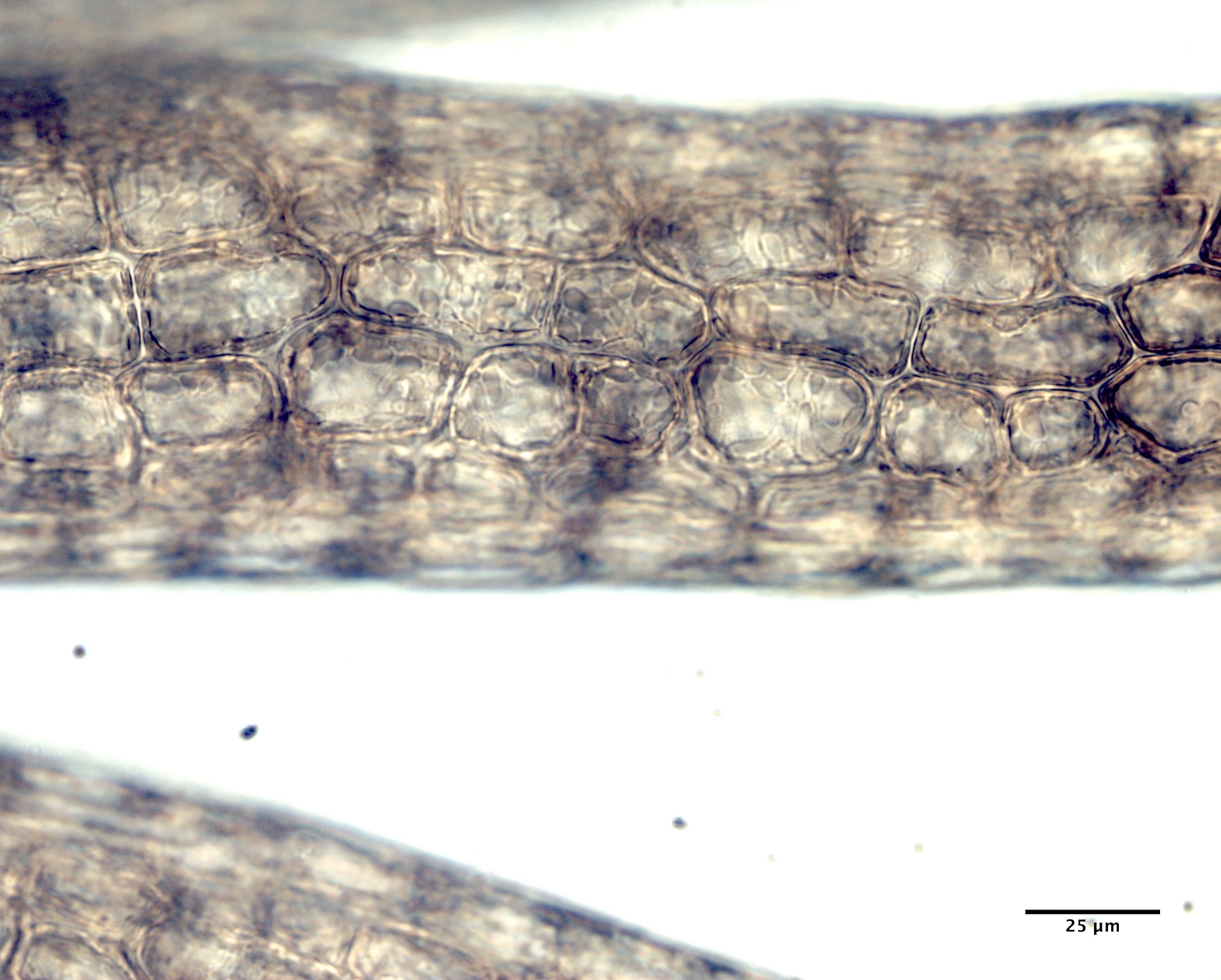 Image F. Cortication masks the central axial filament and polysiphonous construction characteristic of this species (GWS044253).
Image F. Cortication masks the central axial filament and polysiphonous construction characteristic of this species (GWS044253).

Image G. Central axial cell (A) surrounded by six periaxial cells (P) and a thick cortex (GWS044253; mid thallus).

Image H. Central axial cell (A) with associated periaxial cells (P) and similar looking inner cortical cells (C) (subtidal (8 m) on rock, Peggys Cove, NS; GWS011679; near base).

Image I. The actual axial cell (white asterisk; or is it the cell two below this one?) appears to be a periaxial cell in a false rosette the presumptive central cell of which is 100 µm wide, which is not characteristic of this species (low intertidal on rock, Starboard, ME; GWS003665; near base).

Image J. Another section from GWS003665 in which the central axial cell (white asterisk) is unequivocal (near base).

Image K. Young pericarps develop on fertile trichoblasts at the thallus tips (subtidal (2 m) on rock, SW point Mispec Beach, Saint John Harbour monitoring site, NB; EGWS001036).

Image L. Receptive trichogyne extending through an ostiole in the developing pericarp (EGWS001036).

Image M. Spherical cystocarp with terminal ostiole (subtidal (10 m) on rock, Fort Wetherill, RI; GWS006083; rehydrated from press).

Image N. Spherical to oval tetrasporangia formed singly or in pairs (mid intertidal pool on rock; Point Lance (exposed point west of beach), NL; GWS007115; rehydrated from press).
