
Adenophora triphylla var. japonica Inhibits Candida Biofilm Formation, Increases Susceptibility to Antifungal Agents and Reduces Infection


Abstract
:1. Introduction
2. Results
2.1. A. triphylla var. japonica Did Not Inhibit the Fungal Growth
2.2. A. triphylla var. japonica Extract Inhibited the Biofilm Formation of Candida
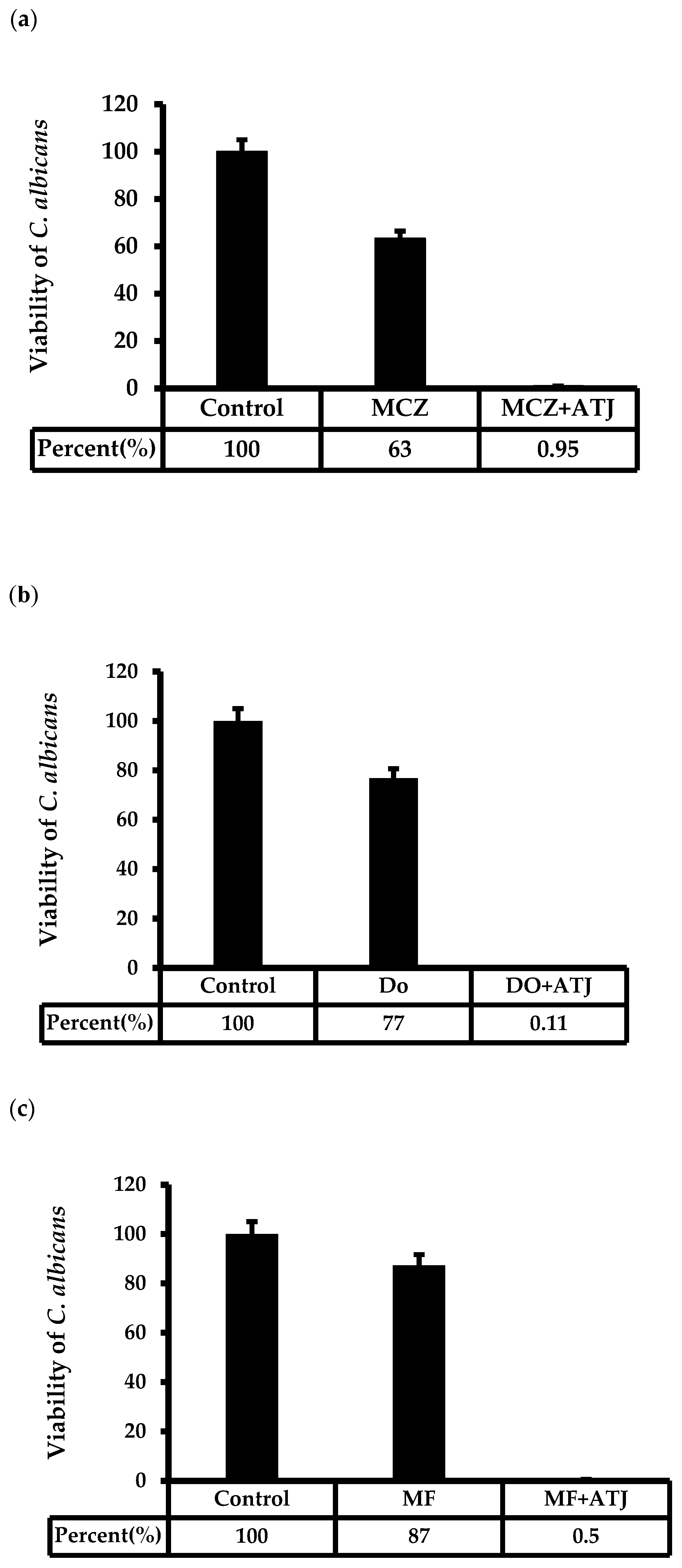
2.3. A. triphylla var. japonica Extract Increased the Susceptibility of Antifungal Agents against C. albicans
2.4. A. triphylla var. japonica Blocked the Dimorphic Transition from Yeast to Hyphae Form
2.5. A. triphylla var. japonica Extract Eeduced Fungal Adherence to the HaCaT Cells
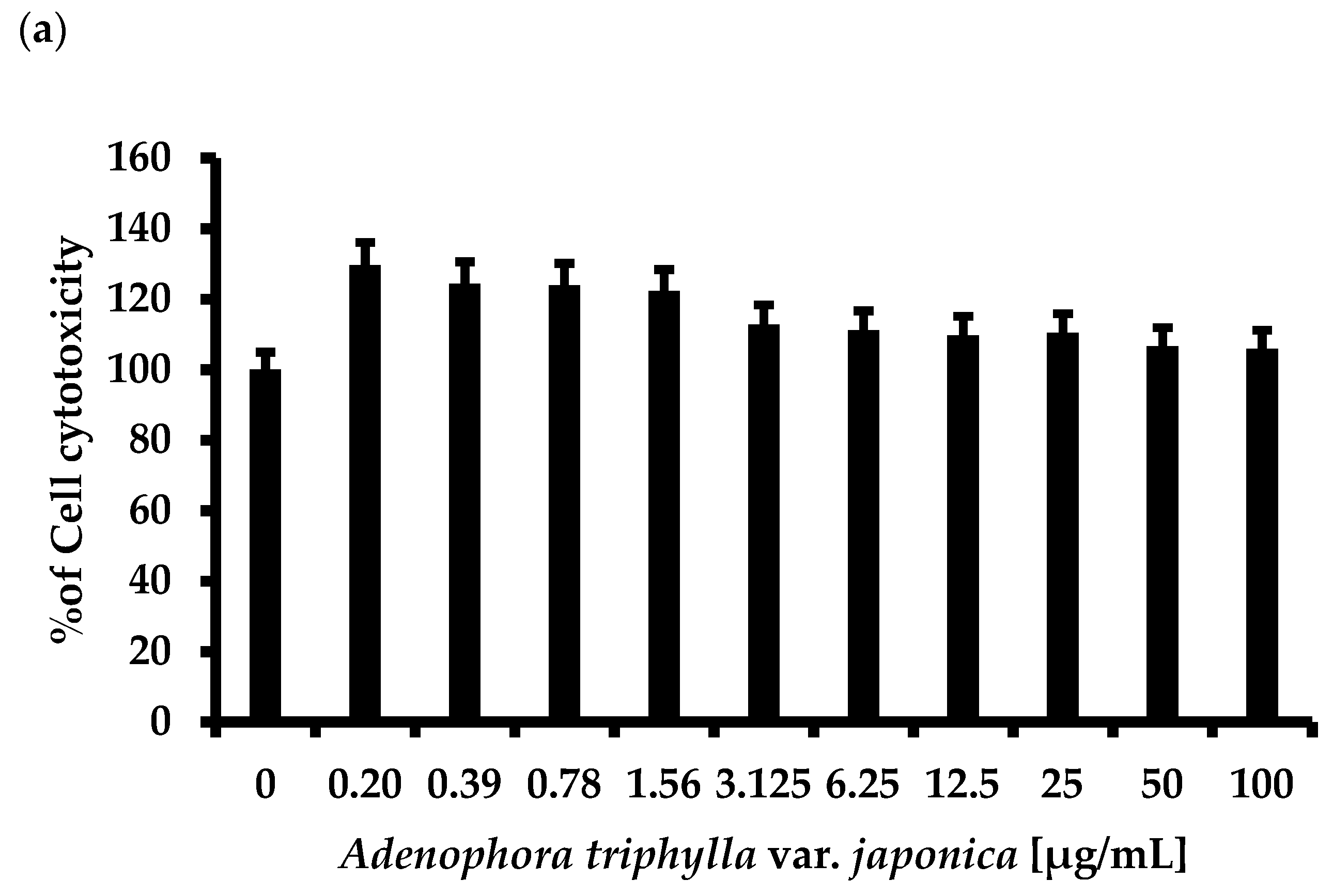
2.6. A. triphylla var. japonica Extract did Not Affect the Growth of Human Originated Cell
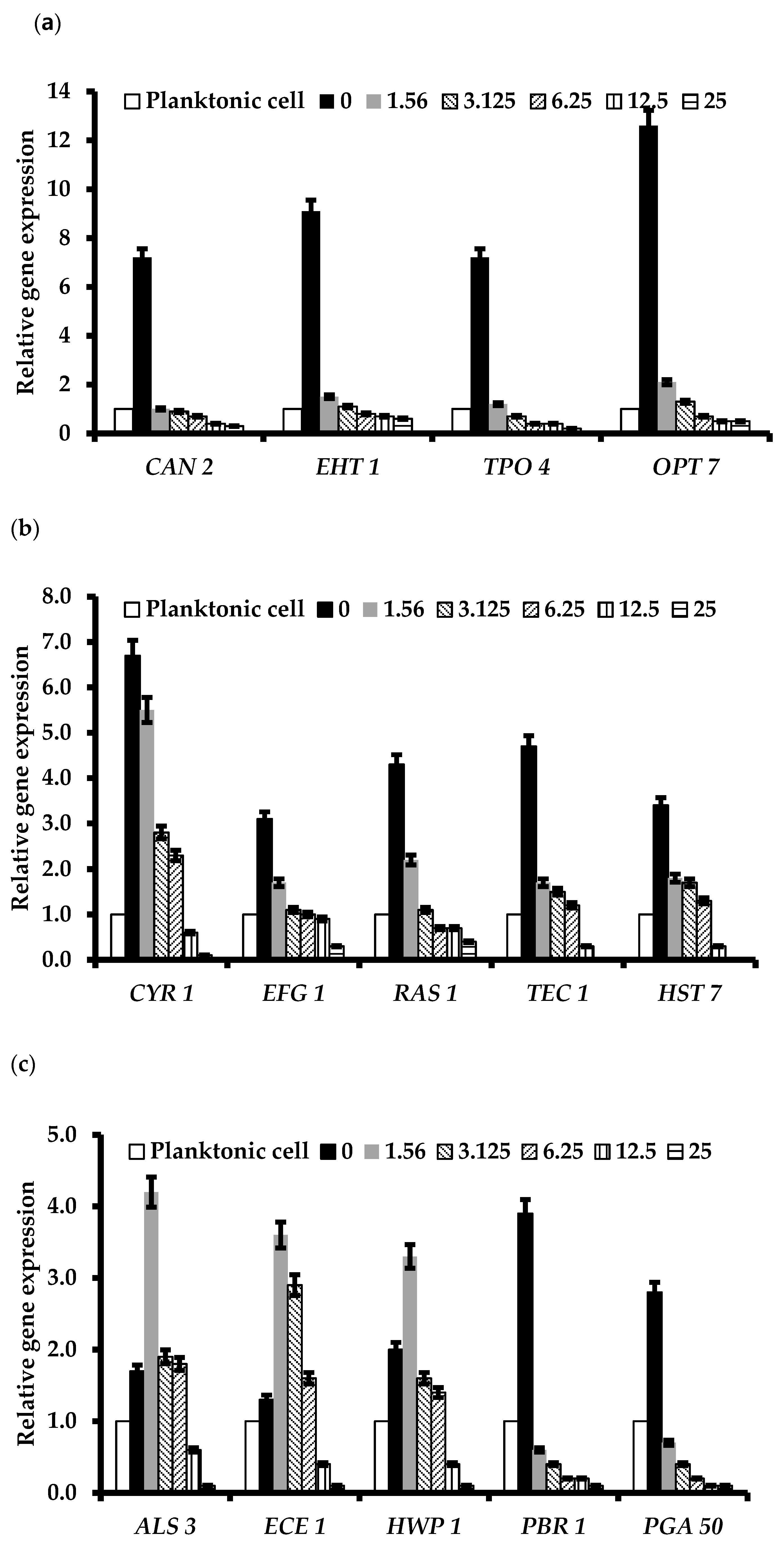
2.7. A. triphylla var. japonica Extract Inhibited the Expression of Biofilm Formation and Infection-Related Genes
3. Discussion
4. Materials and Methods
4.1. Fungal Strains Used in This Study
4.2. Extraction of A. triphylla var. japonica
4.3. Growth Curve Assay
4.4. Biofilm Formation Assay
4.5. Antifungal Activity Assay
4.6. Dimorphic Transition Assay
4.7. Candida Adherence Assay
4.8. MTT Assay
4.9. qRT-PCR Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, F.; Xess, I.; Wang, X.; Jain, N.; Fries, B.C. Biofilm formation in clinical Candida isolates and its association with virulence. Microbes Infect. 2009, 11, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Hogan, D.A.; Sundstrom, P. The Ras/cAMP/PKA signaling pathway and virulence in Candida albicans. Futur. Microbiol. 2009, 4, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Araújo, D.; Henriques, M.; Silva, S. Portrait of Candida Species Biofilm Regulatory Network Genes. Trends Microbiol. 2016, 25, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Van, D.P.; Datta, A. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants of Candida albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Kil, M.; Jung, J.-H.; Kim, J. Roles of Zinc-responsive transcription factor Csr1 in filamentous growth of the pathogenic Yeast Candida albicans. J. Microbiol. Biotechnol. 2008, 18, 242–247. [Google Scholar]
- Lane, S.; Zhou, S.; Pan, T.; Dai, Q.; Liu, H. The Basic Helix-Loop-Helix Transcription Factor Cph2 Regulates Hyphal Development in Candida albicans Partly via Tec1. Mol. Cell. Biol. 2001, 21, 6418–6428. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; López-Ribot, J.L. Candida Biofilms: An Update. Eukaryot. Cell 2005, 4, 633–638. [Google Scholar] [CrossRef] [Green Version]
- Douglas, L. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Ramage, G.; VandeWalle, K.; Lã³Pez-Ribot, J.L.; Wickes, B.L. The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. FEMS Microbiol. Lett. 2002, 214, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, R.P.; Gennings, C. Bloodstream Infections Due to Candida Species in the Intensive Care Unit: Identifying Especially High-Risk Patients to Determine Prevention Strategies. Clin. Infect. Dis. 2005, 41, S389–S393. [Google Scholar] [CrossRef] [PubMed]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm matrix of Candida albicans and Candida tropicalis: Chemical composition and role in drug resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef]
- Siti, N.H.M.A.; Zainuddin, A.M.; Sharifah, R.W.A.; Lee, S.C.; Azizul, A.M.; Nor, A.Y. Herbal processing and extraction technologies. Sep. Purif. Rev. 2016, 45, 305–320. [Google Scholar]
- Nagappan, R. Evaluation of aqueous and ethanol extract of bioactive medicinal plant, Cassia didymobotrya (Fresenius) Irwin & Barneby against immature stages of filarial vector, Culex quinquefasciatus Say (Diptera: Culicidae). Asian Pac. J. Trop. Biomed. 2012, 2, 707–711. [Google Scholar]
- Cota, B.B.; De Oliveira, D.B.C.; Borges, T.C.; Catto, A.C.; Serafim, C.V.; Rodrigues, A.R.A.; Kohlhoff, M.; Zani, C.L.; Andrade, A.A. Antifungal activity of extracts and purified saponins from the rhizomes of Chamaecostus cuspidatus against Candida and Trichophyton species. J. Appl. Microbiol. 2020, 130, 61–75. [Google Scholar] [CrossRef]
- Gorovoi, P.; Ponomarchuk, G.; Strigina, L. A chemotaxonomic study on russian far-eastern campanulaceae. Phytochemistry 1971, 10, 2419–2423. [Google Scholar] [CrossRef]
- Park, S.H. Ethyl Acetate Fraction of Adenophora triphylla var. japonica inhibits migration of lewis lung carcinoma cells by suppressing macrophage polarization toward an M2 phenotype. J. Pharmacopunct. 2019, 22, 253–259. [Google Scholar]
- Chun, J.; Kang, M.; Kim, Y.S. A triterpenoid saponin from Adenophora triphylla var. japonica suppresses the growth of human gastric cancer cells via regulation of apoptosis and autophagy. Tumor Biol. 2014, 35, 12021–12030. [Google Scholar] [CrossRef]
- Ovesna, Z.; Vachalkova, A.; Horvathova, K. Taraxasterol and beta-sitosterol: New naturally compounds with chemoprotective/chemopreventive effects. Neoplasma 2004, 51, 407–414. [Google Scholar]
- Na, M.; Kim, B.Y.; Osada, H.; Ahn, J.S. Inhibition of protein tyrosine phosphatase 1B by lupeol and lupenone isolated from Sorbus commixta. J. Enzym. Inhib. Med. Chem. 2009, 24, 1056–1059. [Google Scholar] [CrossRef]
- Gachet, M.S.; Kunert, O.; Kaiser, M. Antiparasitic compounds from Cupania cinerea with activities against Plasmodium falciparum and Trypanosoma brucei rhodesiense. J. Nat. Prod. 2011, 75, 559–566. [Google Scholar] [CrossRef]
- Cho, J.; Choi, H.; Lee, J.; Kim, M.-S.; Sohn, H.-Y.; Lee, D.G. The antifungal activity and membrane-disruptive action of dioscin extracted from Dioscorea nipponica. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1153–1158. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bao, T.H.Q.; Shin, Y.-K.; Kim, K.-Y. Antifungal activity of magnoflorine against Candida strains. World J. Microbiol. Biotechnol. 2018, 34, 167. [Google Scholar] [CrossRef] [PubMed]
- Lingmei, S.; Kai, L.; Dayoung, W. Effects of magnolol and honokiol on adhesion, yeast-hyphal transition, and formation of biofilm by Candida albicans. PLoS ONE 2015, 10, e0117695. [Google Scholar]
- Khodavandi, A.; Alizadeh, F.; Harmal, N.S.; Sidik, S.M.; Othman, F.; Jahromi, M.A.F.; Sekawi, Z.; Ng, K.-P.; Chong, P.P. Comparison between efficacy of allicin and fluconazole against Candida albicans in vitro and in a systemic candidiasis mouse model. FEMS Microbiol. Lett. 2011, 315, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Paola, M.; Roberta, F.; Cosmeri, R.; Tavanti, A.; Lupetti, A. Inhibition of Candida albicans biofilm formation by the synthetic Lactoferricin derived peptide hLF1-11. PLoS ONE 2016, 11, e0167470. [Google Scholar]
- Zhang, L.L.; Lin, H.; Liu, W.; Dai, B.; Yan, L.; Cao, Y.B.; Jiang, Y.-Y. Antifungal Activity of the ethanol Extract from Flos Rosae Chinensis with activity against Fluconazole-Resistant Clinical Candida. Evid. Based Complement. Altern. Med. 2017, 2017, 4780746. [Google Scholar]
- Bonifácio, B.V.; Vila, T.V.M.; Masiero, I.F.; Silva, P.B.D.; Silva, I.C.D.; Lopes, É.D.O.; dos Santos Ramos, M.A.; Perez de Sousa, L.; Vilegas, W.; Bauab, T.M.; et al. Antifungal activity of a hydroethanolic extract from Astronium urundeuva leaves against Candida albicans and Candida glabrata. Front. Microbiol. 2019, 10, 2642. [Google Scholar] [CrossRef] [Green Version]
- Gacser, A. Adhesins in Candida parapsilosis: Understudied players in virulence. Virulence 2016, 7, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Taff, H.T.; Nett, J.E.; Zarnowski, R.; Ross, K.M.; Sanchez, H.; Cain, M.T.; Hamaker, J.; Mitchell, A.P.; Andes, D.R. A Candida biofilm -induced pathway for matrix glycan delivery: Implications for drug resistance. PLoS Pathog 2012, 8, e1002848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standards, Document M7-A5; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2006. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | IC50 (Half Maximal Inhibitory Concentration) |
|---|---|
| C. albicans | 6.25 µg/mL |
| C. tropicalis var. tropicalis | 25 μg/mL |
| C. parapsilosis var. parapsilosis | 6.25 μg/mL |
| C. glabrata | 6.25 μg/mL |
| C. tropicalis | 25 μg/mL |
| C. parapsilosis | 50 μg/mL |
| Strain | Description | Source |
|---|---|---|
| C. albicans | KCTC 7965 | Purchased form KCTC (Korean Collection for Type of Cultures) or KACC (Korean Agriculture Culture Collection) |
| C. tropicalis var. tropicalis | KCTC17762 | |
| C. parapsilosis var. parapsilosis | KACC45480 | |
| C. glabrata | KCTC7219 | |
| C. tropicalis | KCTC7212 | |
| C. parapsilosis | KACC49573 |
| Genes | Primer Sequence | Gene Function | References |
|---|---|---|---|
| ACT1 | F: TAGGTTTGGAAGCTGCTGG R: CCTGGGAACATGGTAGTAC | Control | [8] |
| CAN2 | F: GCGGAATGGATATGCATGGG R: CGGATTGCTCTTGGAGAAGC | Biofilm formation | In this study |
| EHT1 | F: TCGGAAAGCTTGGTGAAAGC R: ATTTGGCCAAAGCAGGACTC | Biofilm formation | In this study |
| TPO4 | F: GCGGAATGGATATGCATGGG R: CGGATTGCTCTTGGAGAAGC | Biofilm formation | In this study |
| OPT7 | F: TTGATCCCAGCTGCCAAATG R: TGGCCCAAGTTCTTCGTATC | Biofilm formation | In this study |
| CYR1 | F: GTTTCCCCCACCACTCA R: TTGCGGTAATGACACAACAG | Ras1-cAMP-Efg1 pathway | [8] |
| EFG1 | F: TTGAGATGTTGCGGCAGGAT R: ACTGGACAGACAGCAGGAC | Ras1-cAMP-Efg1 pathway | [8] |
| HST7 | F: GCCAGTATGGTCGGAGGAT R: ACATAGGCATCGTCTTCGTC | MAP kinases pathway | [8] |
| RAS1 | F: GAGGTGGTGGTGTTGGTA R: TCTTCTTGTCCAGCAGTATC | Ras1-cAMP-Efg1 pathway | [8] |
| TEC1 | F: GCACTGGCTTCAAGCTCAAA R: GCTGCTGCACCAAGTTCTG | Extracellular matrix | [8] |
| ALS3 | F: GGTTATCGTCCATTTGTTG R: TTCTGTATCCAGTCCATCT | Hyphal-specific genes | [8] |
| ECE1 | F: ACAGTTTCCAGGACGCCAT R: ATTGTTGCTCGTGTTGCCA | Hyphal-specific genes | [8] |
| HWP1 | F: ACAGGTAGACGGTCAAGG R: GGGTAATCATCACATGGTTC | Ras1-cAMP-Efg1 pathway | [8] |
| PBR1 | F: TGTTGCTGCTGGTTCTGATG R: GGTGGCAGATTTGGATTACC | Hyphal-specific genes | [8] |
| PGA50 | F: ATTTCGAAGGGTGCAACTGC R: AAGCACTGCAATGGGAGTTG | Hyphal-specific genes | In this study |
| ADH5 | F: ACCTGCAAGGGCTCATTCTG R: CGGCTCTCAACTTCTCCATA | Extracellular matrix | [8] |
| CSH1 | F: CGTGAGGACGAGAGAGAAT R: CGAATGGACGACACAAAACA | Extracellular matrix | [8] |
| GSC1 | F: CCCATTCTCTAGGCACGA R: ATCAACAACCACTTGCTTCG | Extracellular matrix | [8] |
| ZAP1 | F: ATCTGTCCAGTGTTGTTTGTA R: AGGTCTCTTTGAAAGTTGTG | Extracellular matrix | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Kim, K.-Y. Adenophora triphylla var. japonica Inhibits Candida Biofilm Formation, Increases Susceptibility to Antifungal Agents and Reduces Infection. Int. J. Mol. Sci. 2021, 22, 12523. https://doi.org/10.3390/ijms222212523
Kim D, Kim K-Y. Adenophora triphylla var. japonica Inhibits Candida Biofilm Formation, Increases Susceptibility to Antifungal Agents and Reduces Infection. International Journal of Molecular Sciences. 2021; 22(22):12523. https://doi.org/10.3390/ijms222212523
Chicago/Turabian StyleKim, Daseul, and Ki-Young Kim. 2021. "Adenophora triphylla var. japonica Inhibits Candida Biofilm Formation, Increases Susceptibility to Antifungal Agents and Reduces Infection" International Journal of Molecular Sciences 22, no. 22: 12523. https://doi.org/10.3390/ijms222212523