
Molecular and Morphological Characterization of Colaconema formosanum sp. nov. (Colaconemataceae, Rhodophyta)—A New Endophytic Filamentous Red Algal Species from Taiwan


Abstract
:1. Introduction
2. Materials and Methods
3. Results
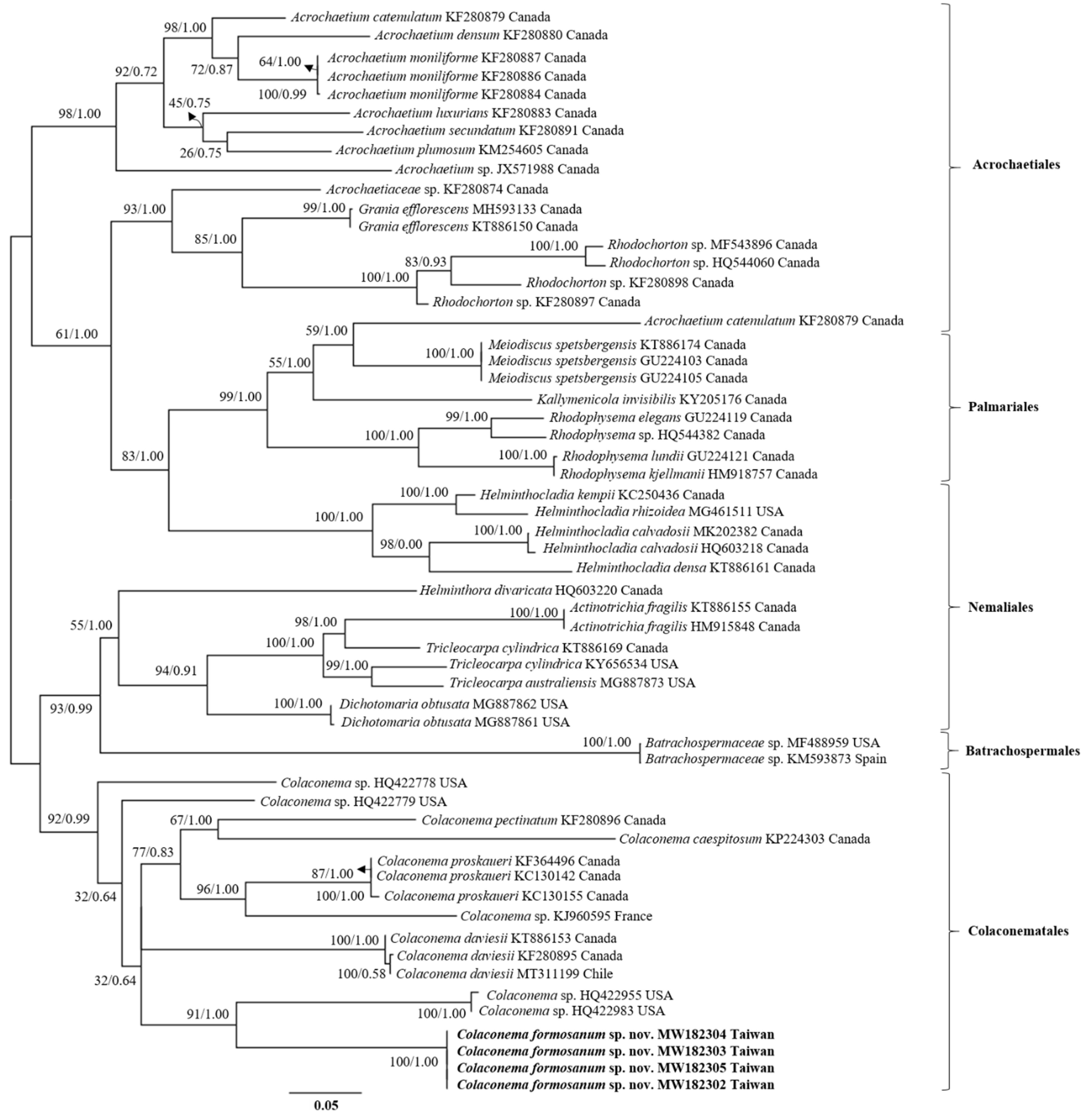
3.1. Phylogenetic Results
3.2. Morphological and Culture Analyses
3.2.1. Vegetative Morphology
3.2.2. Reproductive Morphology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COI-5P | cytochrome c oxidase subunit I gene |
| ML | maximum-likelihood |
| NJ | neighbor-joining |
| PES | Provasoli-enriched seawater |
| rbcL | ribulose-1,5-bisphosphate carboxylase/oxygenase |
References
- Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2021. Available online: https://www.algaebase.org (accessed on 2 March 2021).
- Lam, D.W.; Verbruggen, H.; Saunders, G.W.; Vis, M.L. Multigene phylogeny of the red algal subclass Nemaliophycidae. Mol. Phylogenet. Evol. 2016, 94, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.P.; de Beauclair Guimarães, S.M.P.; Fujii, M.T.; Yoneshigue-Valentin, Y.; Batista, M.G.S.; Yokoya, N.S. Rhodachlya westii sp. nov. (Rhodachlyales, Rhodophyta), a new species from Brazil, revealed by an integrative taxonomic approach. Phycologia 2020, 59, 346–354. [Google Scholar] [CrossRef]
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.; Saunders, G. A molecular systematic investigation of the Acrochaetiales (Florideophycidae, Rhodophyta) and related taxa based on nuclear small-subunit ribosomal DNA sequence data. Eur. J. Phycol. 1998, 33, 221–229. [Google Scholar] [CrossRef]
- Woelkerling, W.J. The Audouinella (Acrochaetium-Rhodochorton) complex (Rhodophyta): Present perspectives. Phycologia 1983, 22, 59–92. [Google Scholar] [CrossRef]
- Harper, J.T.; Saunders, G.W. A re-classification of the Acrochaetiales based on molecular and morphological data, and establishment of the Colaconematales ord. nov. (Florideophyceae, Rhodophyta). Eur. J. Phycol. 2002, 37, 463–476. [Google Scholar] [CrossRef]
- Garbary, D.; Gabrielson, P. Acrochaetiales (Rhodophyta): Taxonomy and evolution. Cryptogam. Algol. 1987, 8, 241–252. [Google Scholar]
- Batters, E.A.L. On some New British Marine Algae. Ann. Bot. 1895, 9, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Saunders, G.W.; Jackson, C.; Salomaki, E. Phylogenetic analyses of transcriptome data resolve familial assignments for genera of the red-algal Acrochaetiales-Palmariales Complex (Nemaliophycidae). Mol. Phylogenet. Evol. 2018, 119, 151–159. [Google Scholar] [CrossRef]
- Skriptsova, A.; Kalita, T. A re-evaluation of Palmaria (Palmariaceae, Rhodophyta) in the North-West Pacific. Eur. J. Phycol. 2020, 55, 266–274. [Google Scholar] [CrossRef]
- Araújo, P.G.; Araújo, P.G.; Schmidt Éder, C.; Kreusch, M.G.; Kano, C.H.; Guimarães, S.M.; Bouzon, Z.L.; Fujii, M.; Yokoya, N.S. Ultrastructural, morphological, and molecular characterization of Colaconema infestans (Colaconematales, Rhodophyta) and its host Kappaphycus alvarezii (Gigartinales, Rhodophyta) cultivated in the Brazilian tropical region. Environ. Boil. Fishes 2014, 26, 1953–1961. [Google Scholar] [CrossRef]
- Montoya, V.; Meynard, A.; Contreras-Porcia, L.; Contador, C.B. Molecular identification, growth, and reproduction of Colaconema daviesii (Rhodophyta; Colaconematales) endophyte of the edible red seaweed Chondracanthus chamissoi. Environ. Boil. Fishes 2020, 32, 3533–3542. [Google Scholar] [CrossRef]
- Wynne, M.; Schneider, C. Colaconema basiramosum sp. nov (Colaconemataceae, Rhodophyta) from the Sultanate of Oman, Northern Arabian Sea. Cryptogam. Algol. 2008, 29, 69–80. [Google Scholar]
- Shih, C.-C.; Hwang, H.-R.; Chang, C.-I.; Su, H.-M.; Chen, P.-C.; Kuo, H.-M.; Li, P.-J.; Tsui, K.-H.; Lin, Y.-C.; Huang, S.-Y.; et al. Anti-Inflammatory and Antinociceptive Effects of Ethyl Acetate Fraction of an Edible Red Macroalgae Sarcodia ceylanica. Int. J. Mol. Sci. 2017, 18, 2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Prieto, C.; De Clerck, O.; Kitayama, T.; Lin, S.-M. Systematic revision of the widespread species Sarcodia ceylanica (Sarcodiaceae, Rhodophyta) in the Indo-Pacific Oceans, including S. suiae sp. nov. Phycologia 2017, 56, 63–76. [Google Scholar] [CrossRef]
- Provasoli, L. Media and Prospects for the Cultivation of Marine Algae. In Proceedings of the US-Japan Conference, Hakone, Japan, 12–15 September 1968; pp. 63–75. [Google Scholar]
- Freshwater, D.W.; Rueness, J. Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 1994, 33, 187–194. [Google Scholar] [CrossRef]
- Lin, S.-M.; Fredericq, S.; Hommersand, M.H. Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on large subunit rDNA and rbcl sequences, including the phycodryoideae, subfam. Nov. J. Phycol. 2001, 37, 881–899. [Google Scholar] [CrossRef] [Green Version]
- Saunders, G.W. Applying DNA barcoding to red macroalgae: A preliminary appraisal holds promise for future applications. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1879–1888. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree. Version 1.4.2 (Inst. Evol. Biol., Univ. Edinburgh, 2014). Available online: http://tree.bio.ed.ac.uk/software/figtree/(accessed on 1 February 2021).
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Hoehna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbary, D. The Effects of Temperature on the Growth and Morphology of Some Audouinella Spp. (Acrochaetiaceae, Rhodophyta). Bot. Mar. 1979, 22, 493–498. [Google Scholar] [CrossRef]
- Tan, P.-L.; Lim, P.-E.; Lin, S.-M.; Phang, S.-M. Halymenia johorensis sp. nov. (Halymeniaceae, Rhodophyta), a new foliose red algal species from Malaysia. Environ. Boil. Fishes 2018, 30, 187–195. [Google Scholar] [CrossRef]
- Boo, G.H.; Kim, K.M. A new species of marine algae from Korea based on morphology and molecular data: Gelidium palmatum sp. nov. (Gelidiales, Rhodophyta). ALGAE 2020, 35, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Bentall, G.B.; Rosen, B.H.; Kunz, J.M.; Miller, M.A.; Saunders, G.W.; Laroche, N.L. Characterization of the putatively introduced red alga Acrochaetium secundatum (Acrochaetiales, Rhodophyta) growing epizoically on the pelage of southern sea otters (Enhydra lutris nereis). Mar. Mammal Sci. 2015, 32, 753–764. [Google Scholar] [CrossRef]
- Andrews, J.H. Pathology of seaweeds: Current status and future prospects. Cell. Mol. Life Sci. 1979, 35, 429. [Google Scholar] [CrossRef]
- Correa, J.; McLachlan, J. Endophytic algae of Chondrus crispus (Rhodophyta). IV Effects on the host following infections by Acrochaete operculata and A. heteroclada (Chlorophyta). Mar. Ecol. Prog. Ser. 1992, 81, 73–87. [Google Scholar] [CrossRef]
- González, M.A.; Goff, L.J. The red algal ekpiphytes microcladia coulteri and m. californica (rhodophyceae, ceramiaceae). J. Phycol. 1989, 25, 558–567. [Google Scholar] [CrossRef]
- Bouzon, Z.L.; Ferreira, E.C.; dos Santos, R.W.; Scherner, F.; Horta, P.A.; Maraschin, M.; Schmidt, E.C. Influences of cadmium on fine structure and metabolism of Hypnea musciformis (Rhodophyta, Gigartinales) cultivated in vitro. Protoplasma 2011, 249, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Chung, C.S.; Hurtado, A.Q.; Soya, F.E.; Lhonneur, G.B.; Critchley, A. Distribution and symptoms of epiphyte infection in major carrageenophyte-producing farms. In Proceedings of the 19th International Seaweed Symposium, Kobe, Japan, 26–31 March 2007; Borowitzka, M.A., Critchley, A.T., Kraan, S., Peters, A., Sjøtun, K., Notoya, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 27–33. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | C. formosanum | C. daviesii | C. infestans |
|---|---|---|---|
| Habit | Marine | Marine | Marine |
| Host | Sarcodia suae | Chondracanthus chamissoi | Kappaphycus alvarezii |
| Thallus long (mm) | 20–250 | - | 5 |
| Cell dimensios (μm) | 6–7.5 × 12–15 | - | 2.4–6.4 × 10–17.6 |
| Cell type | Cylindrical | Cylindrical | Cylindrical |
| Monosporangia | Subspherical, solitary or in clusters | Subspherical | Subspherical |
| Pyrenoid | Single | - | Single |
| Plastid | Single parietal | - | Single parietal |
| Spore (μm) | 9–10 | - | - |
| Spore germination | Type II | Type II | - |
| Phycoerythrin type | R | - | - |
| Type locality | Ping-Tung, Taiwan | Bio-Bio, Chile | Pitimbu, Brazil |
| References | This study | [13] | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-C.; Yeh, H.-Y. Molecular and Morphological Characterization of Colaconema formosanum sp. nov. (Colaconemataceae, Rhodophyta)—A New Endophytic Filamentous Red Algal Species from Taiwan. J. Mar. Sci. Eng. 2021, 9, 809. https://doi.org/10.3390/jmse9080809
Lee M-C, Yeh H-Y. Molecular and Morphological Characterization of Colaconema formosanum sp. nov. (Colaconemataceae, Rhodophyta)—A New Endophytic Filamentous Red Algal Species from Taiwan. Journal of Marine Science and Engineering. 2021; 9(8):809. https://doi.org/10.3390/jmse9080809
Chicago/Turabian StyleLee, Meng-Chou, and Han-Yang Yeh. 2021. "Molecular and Morphological Characterization of Colaconema formosanum sp. nov. (Colaconemataceae, Rhodophyta)—A New Endophytic Filamentous Red Algal Species from Taiwan" Journal of Marine Science and Engineering 9, no. 8: 809. https://doi.org/10.3390/jmse9080809