
Protective Effect of Butanolic Fraction of Delphinium brunonianum on Fructose-Mediated Metabolic Alterations in Rats

 ,
,  , , ,
, , ,  ,
,
Abstract
:
1. Introduction
2. Results
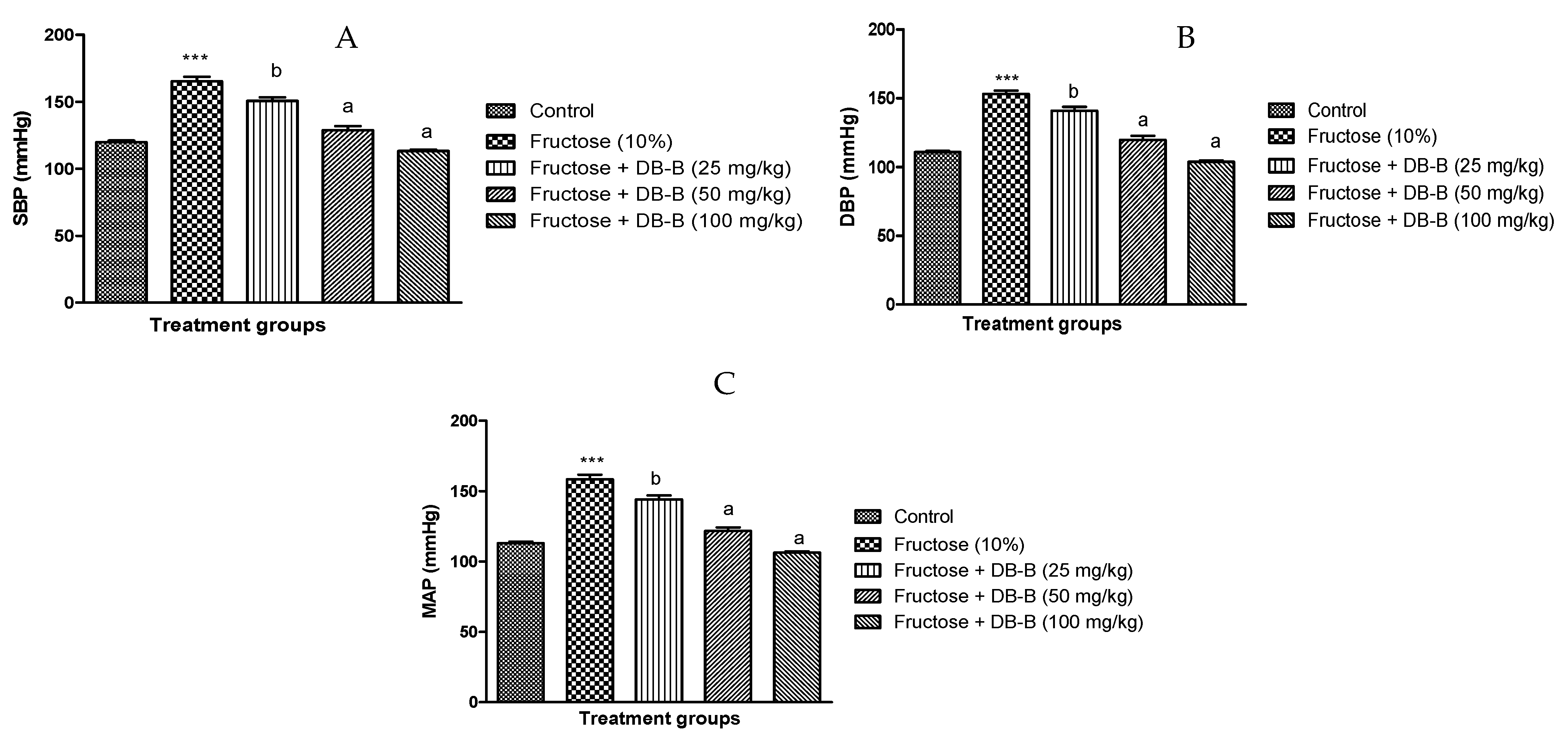
2.1. DB-B-Caused Decrease in Blood Pressure in Fructose-Fed Hypertensive Rats
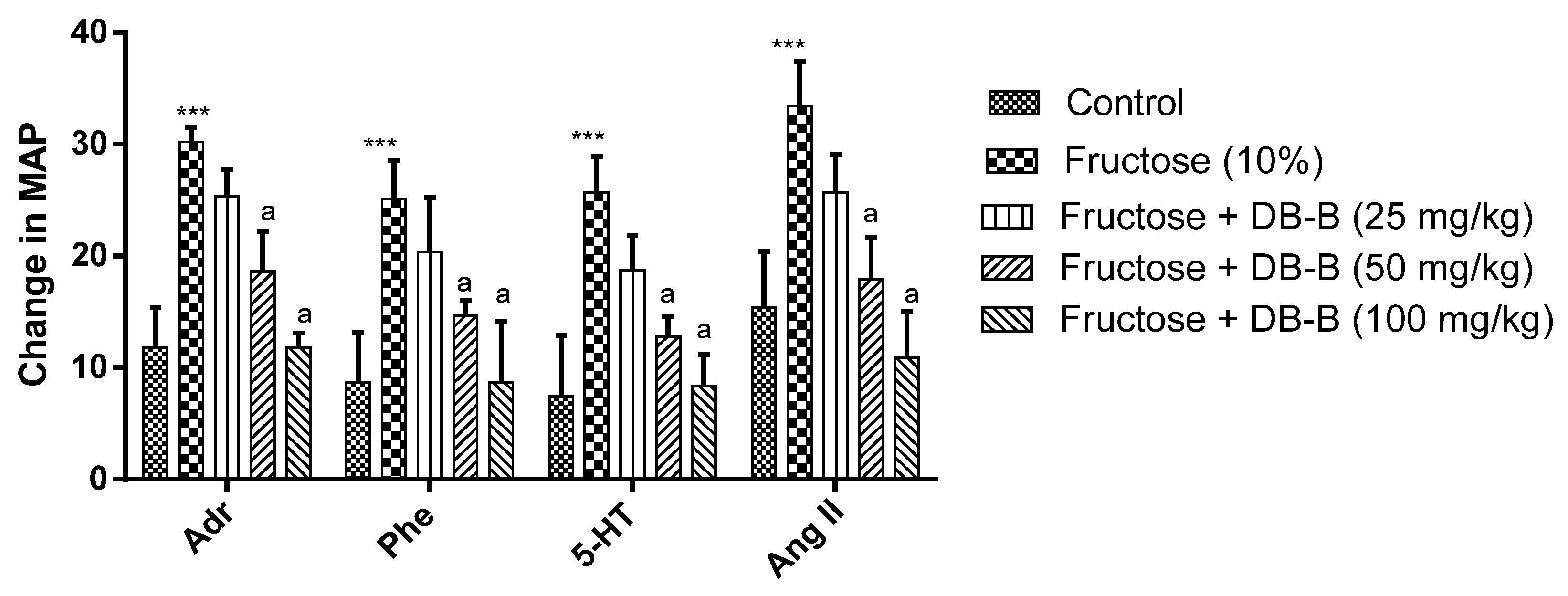
2.2. DB-B-Induced Decrease in Sympathetic Over-Activity in Fructose-Fed Hypertensive Rats
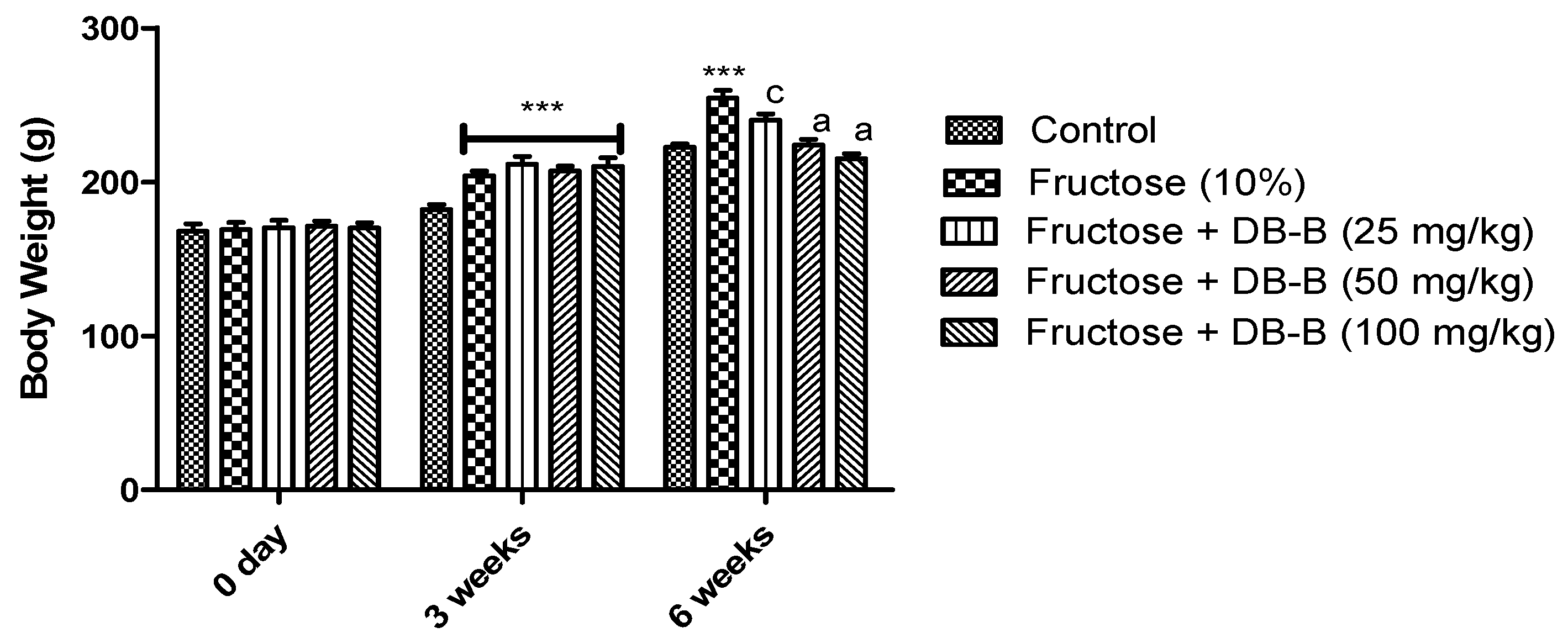
2.3. DB-B-Prevented Increase in Body Weight in Fructose-Fed Rats
2.4. DB-B-Mediated Alteration in Metabolic Abnormalities Triggered by Fructose
2.4.1. Reversal of Fructose-Elicited Dyslipidemia in Rats
2.4.2. Decrease in Level of Uric Acid, Urea, and Creatinine in the Blood of Fructose-Fed Hypertensive Rats
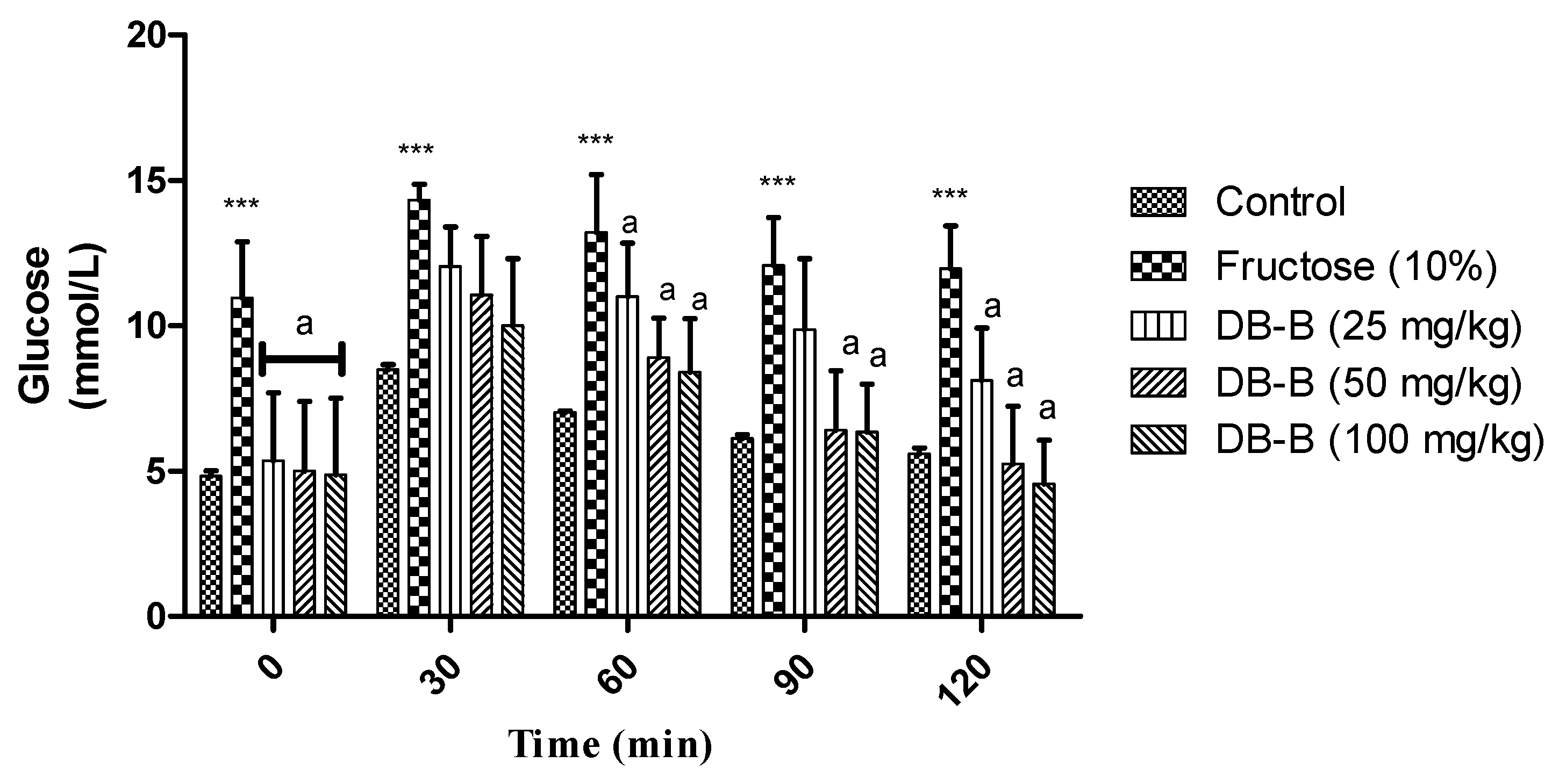
2.4.3. Reversal of Glucose Intolerance and Hyperinsulinemia
2.4.4. D. brunonianum-Evoked Protective Effect on Oxidative Stress Marker in Fructose-Treated Hypertensive Rats
2.4.5. D. brunonianum-Induced Reversal of Acetylcholine-Mediated Vasorelaxation in PE Preconstricted Aortic Tissues
2.5. Phytochemical Analysis of DB-B Using LC-MS
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Chemicals and Drugs Used
4.3. Plant Collection and Preparation of Extract
4.4. Effect of Butanolic Fraction of Delphinium brunonianum on Fructose-Induced Hypertension and Metabolic Alterations in Rats
4.5. Alteration in Fructose-Triggered Metabolic Abnormalities
4.6. Evaluation of DB-B-Mediated Reversal of Glucose Intolerance and Insulin Resistance in Fructose-Fed Rats
4.7. Analysis of Alteration in the Level of Enzymes of the Oxidative System
4.8. Effect of Butanolic Fraction of D. brunonianum on Fructos- Mediated Dysfunction of Vascular Endothelium
4.9. Phytochemical Analysis of DB-B
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldeisen, S.E.; Tucker, K.L. Nutritional strategies in the prevention and treatment of metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 46–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi-Mameghani, M.; Asghari-Jafarabadi, M.; Rezazadeh, K. TCF7L2-rs7903146 polymorphism modulates the effect of artichoke leaf extract supplementation on insulin resistance in metabolic syndrome: A randomized, double-blind, placebo-controlled trial. J. Integr. Med. 2018, 16, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Abdala, S.; Martín-Herrera, D.; Benjumea, D.; Gutiérrez, S.D. Diuretic activity of some Smilax canariensis fractions. J. Ethnopharmacol. 2012, 140, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Mahady, G.B. Global harmonization of herbal health claims. J. Nutr. 2001, 131, 1120S–1123S. [Google Scholar] [CrossRef] [PubMed]
- Kamboj, V.P. Herbal medicine. Curr. Sci. 2000, 78, 35–39. [Google Scholar]
- Samuelsson, G.; Bohlin, L. Drugs of Natural Origin: A Treatise of Pharmacognosy; CRC Press Inc.: Boca Raton, FL, USA, 2017. [Google Scholar]
- Ghimire, S. Medicinal and aromatic plants in the Nepal Himalaya: Status, use, sale and Conservation. Wildlife 1999, 1, 42–52. [Google Scholar]
- Tripathee, H.P.; Sharma, R.P.; Timilsina, Y.P.; Pathak, R.; Devkota, K.P. An assessment of ethnomedicinal use, chemical constituents analysis and bioactivity evaluation on high altitude medicinal plant Delphinium brunonianum of Manang district. Nepal J. Sci. Technol. 2012, 12, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Hassan, M.; Ali, S. Secondary Metabolite Studies of Some Selected Plants of District Gilgit, Gilgit-Baltistan. Int. J. Pharmacogn. Phytochem. Res. 2014, 6, 4. [Google Scholar]
- Asif, H.; Ahmad, M.I.; Alotaibi, N.H.; Alharbi, K.S.; Bukhari, S.N.A.; Saleem, H.; Locatelli, M. Phytochemical analysis and reappraisal of diuretic activity of Delphinium brunonianum Royle and its mode of action in experimental rats. Pak. J. Pharm. Sci. 2020, 33, 1833–1838. [Google Scholar]
- Asif, H.; Alamgeer, A.; Bukhari, I.A.; Vohra, F.; Afzal, S.; Khan, S.W.; Niazi, Z.R. Phytochemical analysis of crude extract of Delphinium brunonianum and its effect on hypertension and metabolic perturbations in fructose fed rats. Nat. Prod. Res. 2019, 5, 2982–2986. [Google Scholar] [CrossRef]
- Verma, S.; Bhanot, S.; Mc Neill, J.H. Sympathectomy prevents fructose-induced hyperinsulinemia and hypertension. Eur. J. Pharmacol. 1999, 373, R1–R4. [Google Scholar] [CrossRef]
- Tran, L.T.; Yuen, V.G.; McNeill, J.H. The fructose-fed rat: A review on the mechanisms of fructose-induced insulin resistance and hypertension. Mol. Cell. Biochem. 2009, 332, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Elkayam, A.; Mirelman, D.; Peleg, E.; Wilchek, M.; Miron, T.; Rabinkov, A.; Oron-Herman, M.; Rosenthal, T. The effects of allicin on weight in fructose-induced hyperinsulinemic, hyperlipidemic, hypertensive rats. Am. J. Hypertens. 2003, 16, 1053–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahangarpour, A.; Mohammadian, M.; Dianat, M. Antidiabetic effect of hydroalcholic Urtica dioica leaf extract in male rats with fructose-induced insulin resistance. Iran. J. Med. Sci. 2012, 37, 181. [Google Scholar]
- Vikrant, V.; Grover, J.; Tandon, N.; Rathi, S.; Gupta, N. Treatment with extracts of Momordica charantia and Eugenia jambolana prevents hyperglycemia and hyperinsulinemia in fructose fed rats. J. Ethnopharmacol. 2001, 76, 139–143. [Google Scholar] [CrossRef]
- Dimo, T.; Azay, J.; Tan, P.V.; Pellecuer, J.; Cros, G.; Bopelet, M.; Serrano, J.J. Effects of the aqueous and methylene chloride extracts of Bidens pilosa leaf on fructose-hypertensive rats. J. Ethnopharmacol. 2001, 76, 215–221. [Google Scholar] [CrossRef]
- Dimo, T.; Rakotonirina, S.V.; Tan, P.V.; Azay, J.; Dongo, E.; Cros, G. Leaf methanol extract of Bidens pilosa prevents and attenuates the hypertension induced by high-fructose diet in Wistar rats. J. Ethnopharmacol. 2002, 83, 183–191. [Google Scholar] [CrossRef]
- Rösen, P.; Ohly, P.; Gleichmann, H. Experimental benefit of moxonidine on glucose metabolism and insulin secretion in the fructose-fed rat. J. Hypertens. 1997, 15, S31–S38. [Google Scholar] [CrossRef]
- Bantle, J.P. Dietary fructose and metabolic syndrome and diabetes. J. Nutr. 2009, 139, 1263S–1268S. [Google Scholar] [CrossRef] [Green Version]
- Softic, S.; Cohen, D.E.; Kahn, C.R. Role of dietary fructose and hepatic de novo lipogenesis in fatty liver disease. Dig. Dis. Sci. 2016, 61, 1282–1293. [Google Scholar] [CrossRef] [Green Version]
- Zang, L.-Y.; Cosma, G.; Gardner, H.; Shi, X.; Castranova, V.; Vallyathan, V. Effect of antioxidant protection by p-coumaric acid on low-density lipoprotein cholesterol oxidation. Am. J. Physiol. Cell Physiol. 2000, 279, C954–C960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Tuttle, K.R.; Short, R.A.; Johnson, R.J. Hypothesis: Fructose-induced hyperuricemia as a causal mechanism for the epidemic of the metabolic syndrome. Nat. Rev. Nephrol. 2005, 1, 80. [Google Scholar] [CrossRef] [PubMed]
- Feig, D.I. Hyperuricemia and hypertension. Adv. Chronic Kidney Dis. 2012, 19, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Khosla, U.M.; Zharikov, S.; Finch, J.L.; Nakagawa, T.; Roncal, C.; Mu, W.; Krotova, K.; Block, E.R.; Prabhakar, S.; Johnson, R.J. Hyperuricemia induces endothelial dysfunction. Kidney Int. 2005, 67, 1739–1742. [Google Scholar] [CrossRef] [Green Version]
- Olatunji, L.A.; Okwusidi, J.I.; Soladoye, A.O. Antidiabetic effect of Anacardium occidentale. Stem-bark in fructose-diabetic rats. Pharm. Biol. 2005, 43, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Kamalakkannan, N.; Prince, P.S.M. Antihyperglycaemic and antioxidant effect of rutin, a polyphenolic flavonoid, in streptozotocin-induced diabetic wistar rats. Basic Clin. Pharmacol. Toxicol. 2006, 98, 97–103. [Google Scholar] [CrossRef]
- Sánchez-Lozada, L.G.; Tapia, E.; Jiménez, A.; Bautista, P.; Cristóbal, M.; Nepomuceno, T.; Soto, V.; Ávila-Casado, C.; Nakagawa, T.; Johnson, R.J. Fructose-induced metabolic syndrome is associated with glomerular hypertension and renal microvascular damage in rats. Am. J. Physiol. Ren. Physiol. 2007, 292, F423–F429. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Wong, W.-Y.; Kauter, K.; Ward, L.C.; Brown, L. Caffeine attenuates metabolic syndrome in diet-induced obese rats. Nutrition 2012, 28, 1055–1062. [Google Scholar] [CrossRef]
- Verma, S.; Bhanot, S.; Yao, L.; McNeill, J.H. Defective endothelium-dependent relaxation in fructose-hypertensive rats. Am. J. Hypertens. 1996, 9, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Deedwania, P.C. Mechanisms of endothelial dysfunction in the metabolic syndrome. Curr. Diabetes Rep. 2003, 3, 289–292. [Google Scholar] [CrossRef]
- May, J.M. How does ascorbic acid prevent endothelial dysfunction? Free Radic. Biol. Med. 2000, 28, 1421–1429. [Google Scholar] [CrossRef]
- Ren, B.; Qin, W.; Wu, F.; Wang, S.; Pan, C.; Wang, L.; Zeng, B.; Ma, S.; Liang, J. Apigenin and naringenin regulate glucose and lipid metabolism, and ameliorate vascular dysfunction in type 2 diabetic rats. Eur. J. Pharmacol. 2016, 773, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P. Endothelial dysfunction in cardiovascular disease. In Proceedings of the Southeast Asian-Western Pacific Regional Meeting of Pharmacologists, Busan, Korea, 19–23 August 2003. [Google Scholar]
- Hecker, P.A.; Galvao, T.F.; O’Shea, K.M.; Brown, B.H.; Henderson Jr, R.; Riggle, H.; Gupte, S.A.; Stanley, W.C. High-sugar intake does not exacerbate metabolic abnormalities or cardiac dysfunction in genetic cardiomyopathy. Nutrition 2012, 28, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, P.-W.; Lee, H.-C.; Lu, P.-J.; Chen, H.-H.; Lai, C.-C.; Sun, G.-C.; Yeh, T.-C.; Hsiao, M.; Lin, Y.-T.; Liu, C.-P. Resveratrol inhibition of Rac1-derived reactive oxygen species by AMPK decreases blood pressure in a fructose-induced rat model of hypertension. Sci. Rep. 2016, 6, 25342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Tu, Y.-C.; Lian, T.-W.; Hung, J.-T.; Yen, J.-H.; Wu, M.-J. Distinctive antioxidant and antiinflammatory effects of flavonols. J. Agric. Food Chem. 2006, 54, 9798–9804. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, M.; El-Mahdy, M.A.; Abd-Ellah, M.F.; Helal, G.; Khalifa, F.; Hamada, F. Influence of p-coumaric acid on doxorubicin-induced oxidative stress in rat’s heart. Pharmacol. Res. 2003, 48, 461–465. [Google Scholar] [CrossRef]
- De Sousa, E.; Zanatta, L.; Seifriz, I.; Creczynski-Pasa, T.B.; Pizzolatti, M.G.; Szpoganicz, B.; Silva, F.R.M.B. Hypoglycemic Effect and Antioxidant Potential of Kaempferol-3, 7-O-(α)-dirhamnoside from Bauhinia f orficata Leaves. J. Nat. Prod. 2004, 67, 829–832. [Google Scholar] [CrossRef]
- Yen, G.-C.; Duh, P.-D.; Tsai, H.-L. Antioxidant and pro-oxidant properties of ascorbic acid and gallic acid. Food Chem. 2002, 79, 307–313. [Google Scholar] [CrossRef]
- Hung, T.M.; Na, M.; Thuong, P.T.; Su, N.D.; Sok, D.; Song, K.S.; Seong, Y.H.; Bae, K. Antioxidant activity of caffeoyl quinic acid derivatives from the roots of Dipsacus asper Wall. J. Ethnopharmacol. 2006, 108, 188–192. [Google Scholar] [CrossRef]
- Khan, H.; Amin, H.; Ullah, A.; Saba, S.; Rafique, J.; Khan, K.; Ahmad, N.; Badshah, S.L. Antioxidant and antiplasmodial activities of bergenin and 11-O-galloylbergenin isolated from Mallotus philippensis. Oxidative Med. Cell. Longev. 2016, 2016, 1051925. [Google Scholar] [CrossRef] [Green Version]
- Nafisi, S.; Hashemi, M.; Rajabi, M.; Tajmir-Riahi, H.A. DNA adducts with antioxidant flavonoids: Morin, apigenin, and naringin. DNA Cell Biol. 2008, 27, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bellamkonda, R.; Karuna, R.; Rao, B.S.B.; Haritha, K.; Manjunatha, B.; Silpa, S.; Saralakumari, D. Beneficiary effect of Commiphora mukul ethanolic extract against high fructose diet induced abnormalities in carbohydrate and lipid metabolism in wistar rats. J. Tradit. Complementary Med. 2018, 8, 203–211. [Google Scholar] [CrossRef]
- De AF Da, R.D.C.; de Souza, P.; Crestani, S.; Júnior, A.G.; Boligon, A.A.; Athayde, M.L.; da Silva-Santos, J.E. Hypotensive and diuretic effect of the butanolic soluble fraction of the hydroethanolic extract of bark of Scutia buxifolia Reissek in rats. J. Ethnopharmacol. 2015, 172, 395–401. [Google Scholar]
- Mohan, M.; Jaiswal, B.S.; Kasture, S. Effect of Solanum torvum on blood pressure and metabolic alterations in fructose hypertensive rats. J. Ethnopharmacol. 2009, 126, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Moraes, É.A.; da Silva Marineli, R.; Lenquiste, S.A.; Queiroz, V.A.V.; Camargo, R.L.; Borck, P.C.; Carneiro, E.M.; Júnior, M.R.M. Whole sorghum flour improves glucose tolerance, insulin resistance and preserved pancreatic islets function in obesity diet-induced rats. J. Funct. Foods 2018, 45, 530–540. [Google Scholar] [CrossRef]
- Younis, W.; Alamgeer; Schini-Kerth., V.B.; Junior, A.G.; Muhammad, M. Cardioprotective effect of Asphodelus tenuifolius Cav. on blood pressure and metabolic alterations in glucose-induced metabolic syndrome rats–An ethnopharmacological approach. J. Ethnopharmacol. 2018, 214, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Maneesai, P.; Bunbupha, S.; Kukongviriyapan, U.; Prachaney, P.; Tangsucharit, P.; Kukongviriyapan, V.; Pakdeechote, P. Asiatic acid attenuates renin-angiotensin system activation and improves vascular function in high-carbohydrate, high-fat diet fed rats. BMC Complementary Altern. Med. 2016, 16, 123. [Google Scholar] [CrossRef] [Green Version]
- Saleem, H.; Htar, T.T.; Naidu, R.; Nawawi, N.S.; Ahmad, I.; Ashraf, M.; Ahemad, N. Biological, chemical and toxicological perspectives on aerial and roots of Filago germanica (L.) huds: Functional approaches for novel phyto-pharmaceuticals. Food Chem. Toxicol. 2019, 123, 363–373. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.NO | RT (min) | Base Peak m/z | Peak Height | Proposed Compound | Molecular Formula | Molecular Mass | Volume |
|---|---|---|---|---|---|---|---|
| 1 | 2.652 | 217.0481 | 46,409 | bis(4-fluorophenyl)-Methanone | C13 H8 F2 O | 218.0534 | 187,240 |
| 2 | 2.668 | 215.0321 | 5966 | 2-C-Methyl-D-erythritol 4-phosphate | C5 H13 O7 P | 216.0395 | 24,028 |
| 3 | 2.672 | 181.0712 | 18,085 | D-Sorbitol | C6 H14 O6 | 182.0785 | 76,215 |
| 4 | 2.697 | 179.0557 | 9855 | L-Galactose | C6 H12 O6 | 180.0631 | 48,970 |
| 5 | 2.753 | 135.0299 | 8636 | D-threonic acid | C4 H8 O5 | 136.0371 | 40,365 |
| 6 | 2.972 | 366.1153 | 12,110 | Met Ser Met | C13 H25 N3 O5 S2 | 367.1229 | 76,009 |
| 7 | 3.214 | 133.0147 | 48,234 | D-(+)-Malic acid | C4 H6 O5 | 134.022 | 233,663 |
| 8 | 3.57 | 290.0883 | 7253 | Sarmentosin epoxide | C11 H17 N O8 | 291.0956 | 75,515 |
| 9 | 3.832 | 253.0929 | 7434 | Galactosylglycerol | C9 H18 O8 | 254.1001 | 46,868 |
| 10 | 4.028 | 128.0356 | 34,200 | N-Acryloylglycine | C5 H7 N O3 | 129.0429 | 208,646 |
| 11 | 4.152 | 243.0619 | 7285 | Uridine | C9 H12 N2 O6 | 244.0691 | 49,247 |
| 12 | 4.292 | 188.0562 | 8154 | Glutarylglycine | C7 H11 N O5 | 189.0635 | 51,869 |
| 13 | 4.292 | 129.0198 | 6037 | Glutaconic acid | C5 H6 O4 | 130.027 | 39,902 |
| 14 | 4.293 | 173.0091 | 14,206 | Dehydroascorbic acid | C6 H6 O6 | 174.0164 | 91,494 |
| 15 | 4.995 | 117.0195 | 20,991 | Erythrono-1,4-lactone | C4 H6 O4 | 118.0268 | 139,568 |
| 16 | 5.681 | 129.0194 | 6310 | Glutaconic acid | C5 H6 O4 | 130.0267 | 50,742 |
| 17 | 9.83 | 131.0355 | 11,447 | 3-Hydroxy-3-methyl-2-oxo-Butyric acid | C5 H8 O4 | 132.0428 | 58,944 |
| 18 | 10.632 | 293.1244 | 10,448 | Ethyl (S)-3-hydroxybutyrate glucoside | C12 H22 O8 | 294.1317 | 43,798 |
| 19 | 10.732 | 327.0723 | 17,123 | Bergenin | C14 H16 O9 | 328.0795 | 84,427 |
| 20 | 11.07 | 353.0885 | 54,518 | Scopolin | C16 H18 O9 | 354.0957 | 369,833 |
| 21 | 11.071 | 467.0816 | 6864 | Castamollissin | C20 H20 O13 | 468.0887 | 34,751 |
| 22 | 11.277 | 609.1477 | 130,491 | Robinetin 3-rutinoside | C27 H30 O16 | 610.1547 | 1,136,964 |
| 23 | 11.283 | 175.0611 | 6144 | 3-propylmalic acid | C7 H12 O5 | 176.0685 | 28,288 |
| 24 | 11.437 | 340.1558 | 11,223 | Carboxyterbinafine derivative2 | C20 H23 N O4 | 341.163 | 68,196 |
| 25 | 11.437 | 403.151 | 10,458 | Desmethylnimodipine | C20 H24 N2 O7 | 404.1586 | 55,189 |
| 26 | 11.565 | 593.1513 | 47,204 | Luteolin 7-rhamnosyl(1- > 6)galactoside | C27 H30 O15 | 594.1586 | 266,559 |
| 27 | 11.662 | 651.157 | 105,640 | Kaempferol 3-(6″-acetylglucoside)-7-glucoside | C29 H32 O17 | 652.1642 | 849,836 |
| 28 | 11.78 | 609.1452 | 15,662 | Robinetin 3-rutinoside | C27 H30 O16 | 610.1524 | 90,280 |
| 29 | 12.016 | 295.0453 | 38,925 | Mono-trans-p-coumaroylmesotartaric acid | C13 H12 O8 | 296.0527 | 394,426 |
| 30 | 12.051 | 635.1622 | 25,444 | Fujikinetin 7-O-laminaribioside | C29 H32 O16 | 636.1699 | 152,900 |
| 31 | 12.052 | 651.1574 | 86,304 | Kaempferol 3-(6″-acetylglucoside)-7-glucoside | C29 H32 O17 | 652.1647 | 852,487 |
| 32 | 12.089 | 186.1143 | 12,154 | KAPA | C9 H17 N O3 | 187.1216 | 70,741 |
| 33 | 12.231 | 593.1528 | 5324 | Luteolin 7-rhamnosyl(1- > 6)galactoside | C27 H30 O15 | 594.16 | 28,075 |
| 34 | 12.254 | 463.0881 | 19,178 | 8-Hydroxyluteolin 8-glucoside | C21 H20 O12 | 464.0951 | 108,984 |
| 35 | 12.336 | 635.1623 | 13,934 | Fujikinetin 7-O-laminaribioside | C29 H32 O16 | 636.1698 | 78,444 |
| 36 | 12.448 | 635.1623 | 9397 | Fujikinetin 7-O-laminaribioside | C29 H32 O16 | 636.1692 | 52,438 |
| 37 | 12.473 | 693.1673 | 49,263 | Isoscutellarein 7-(6‴-acetylallosyl-(1- > 2)-6″-acetylglucoside) | C31 H34 O18 | 694.1745 | 405,976 |
| 38 | 12.478 | 555.1977 | 5318 | punaglandin 1 | C27 H37 Cl O10 | 556.2062 | 33,134 |
| 39 | 12.62 | 693.1672 | 41,258 | Isoscutellarein 7-(6″-acetylallosyl-(1- > 2)-6″-acetylglucoside) | C31 H34 O18 | 694.1743 | 287,244 |
| 40 | 12.717 | 597.2099 | 8960 | 12S-acetoxy-punaglandin 1 | C29 H39 Cl O11 | 598.2179 | 47,677 |
| 41 | 12.766 | 163.0399 | 7568 | m-Coumaric acid | C9 H8 O3 | 164.047 | 33,265 |
| 42 | 12.859 | 677.171 | 11,272 | Apigenin 7-(4″,6″-diacetylalloside)-4′-alloside | C31 H34 O17 | 678.1785 | 66,085 |
| 43 | 13.133 | 677.1739 | 7170 | Apigenin 7-(4″,6″-diacetylalloside)-4′-alloside | C31 H34 O17 | 678.1804 | 40,991 |
| 44 | 13.137 | 447.0937 | 11,848 | 6-Hydroxyluteolin 5-rhamnoside | C21 H20 O11 | 448.1007 | 70,221 |
| 45 | 13.431 | 121.029 | 6423 | 3-Hydroxybenzaldehyde | C7 H6 O2 | 122.0362 | 28,930 |
| 46 | 13.437 | 237.0393 | 6940 | Dipyrocetyl | C11 H10 O6 | 238.0465 | 31,193 |
| 47 | 13.438 | 699.3497 | 96,124 | Septentriodine | C37 H52 N2 O11 | 700.3569 | 634,267 |
| 48 | 13.705 | 269.1032 | 7341 | Idebenone Metabolite (Benzenebutanoic acid,2,5-dihydroxy-3,4-dimethoxy-6-methyl-) | C13 H18 O6 | 270.1104 | 40,569 |
| 49 | 16.669 | 221.1184 | 34,676 | (6S)-dehydrovomifoliol | C13 H18 O3 | 222.1257 | 211,059 |
| 50 | 17.71 | 293.1769 | 12,618 | Gingerol | C17 H26 O4 | 294.1842 | 81,405 |
| 51 | 18.219 | 309.1711 | 10,585 | methyl 8-[2-(2-formyl-vinyl)-3-hydroxy-5-oxo-cyclopentyl]-octanoate | C17 H26 O5 | 310.1785 | 70,372 |
| 52 | 19.584 | 277.1809 | 7965 | 6-Paradol | C17 H26 O3 | 278.1881 | 50,566 |
| 53 | 21.808 | 265.1489 | 22,964 | Lauryl hydrogen sulfate | C12 H26 O4 S | 266.1563 | 362,120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bukhari, S.N.A.; Asif, H.; Alamgeer; Asim, M.H.; Muhammad Irfan, H.; Ejaz, H.; Elsherif, M.A.; Junaid, K. Protective Effect of Butanolic Fraction of Delphinium brunonianum on Fructose-Mediated Metabolic Alterations in Rats. Metabolites 2022, 12, 481. https://doi.org/10.3390/metabo12060481
Bukhari SNA, Asif H, Alamgeer, Asim MH, Muhammad Irfan H, Ejaz H, Elsherif MA, Junaid K. Protective Effect of Butanolic Fraction of Delphinium brunonianum on Fructose-Mediated Metabolic Alterations in Rats. Metabolites. 2022; 12(6):481. https://doi.org/10.3390/metabo12060481
Chicago/Turabian StyleBukhari, Syed Nasir Abbas, Hira Asif, Alamgeer, Mulazim Hussain Asim, Hafiz Muhammad Irfan, Hasan Ejaz, Mervat A. Elsherif, and Kashaf Junaid. 2022. "Protective Effect of Butanolic Fraction of Delphinium brunonianum on Fructose-Mediated Metabolic Alterations in Rats" Metabolites 12, no. 6: 481. https://doi.org/10.3390/metabo12060481