
Salicornia europaea L. Functional Traits Indicate Its Optimum Growth

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
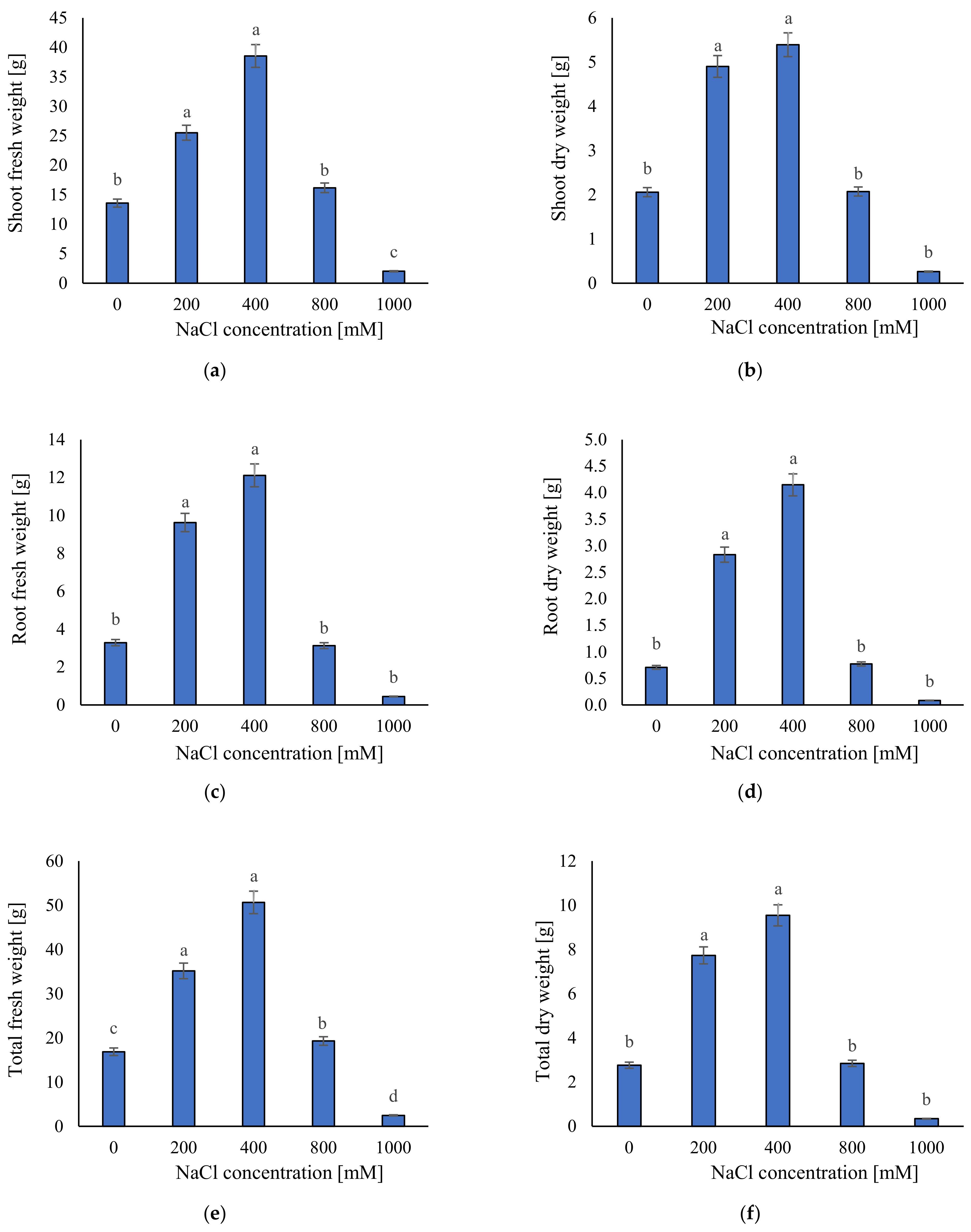
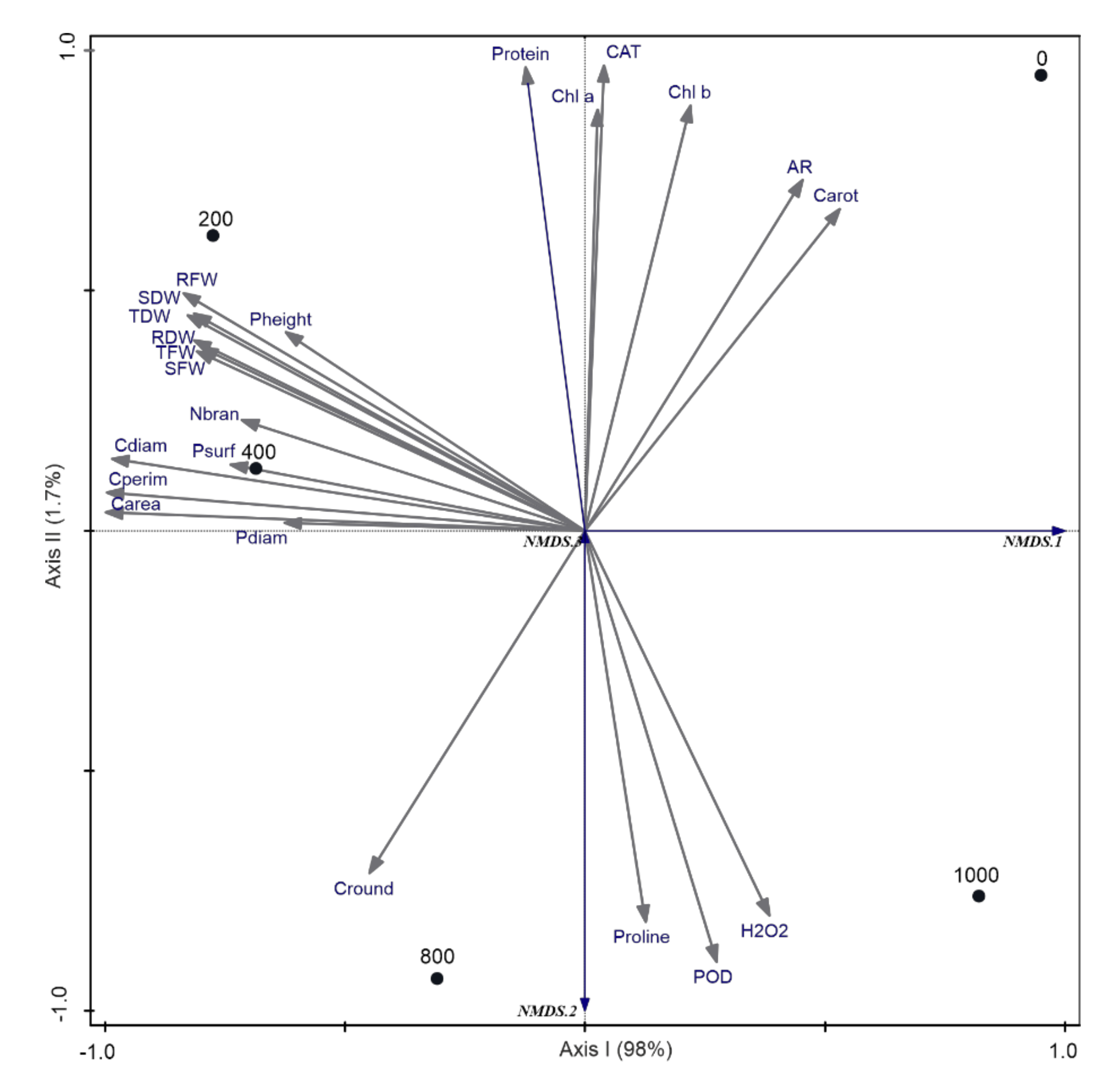
2.1. Growth Responses to Different Salinity Levels
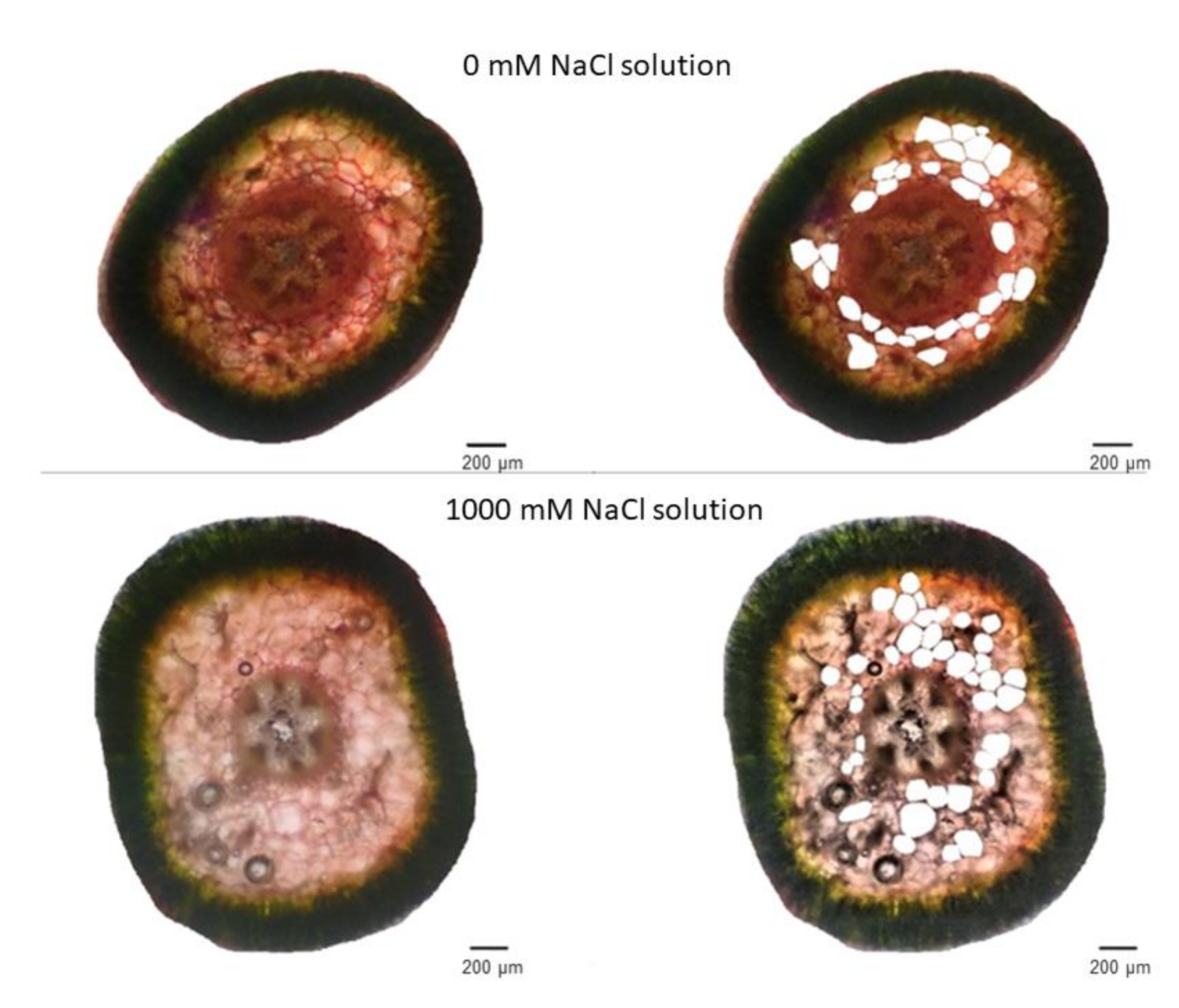
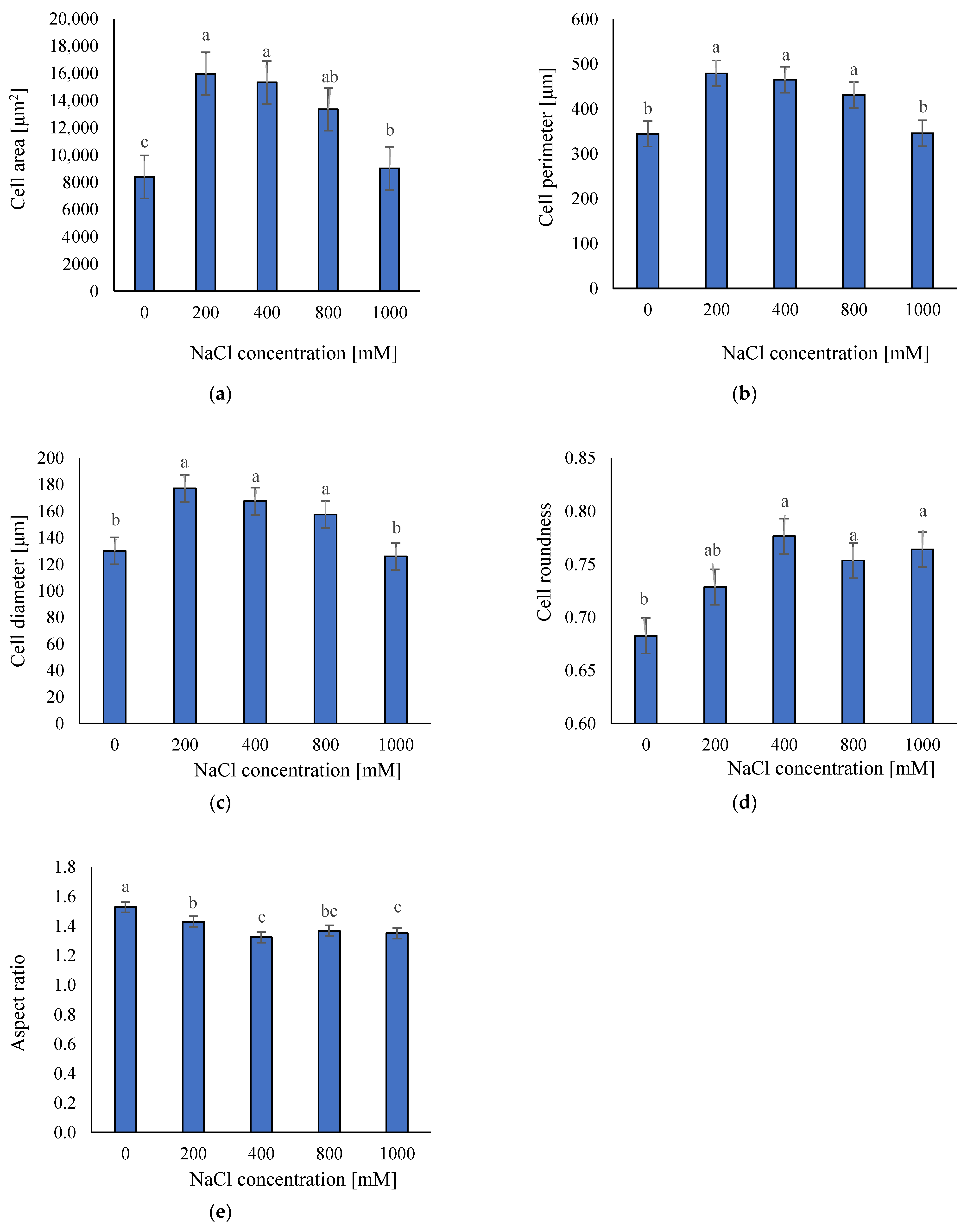
2.2. Anatomical Responses to Different Salinity Levels
2.3. Biochemical Response to Different Salinity Levels
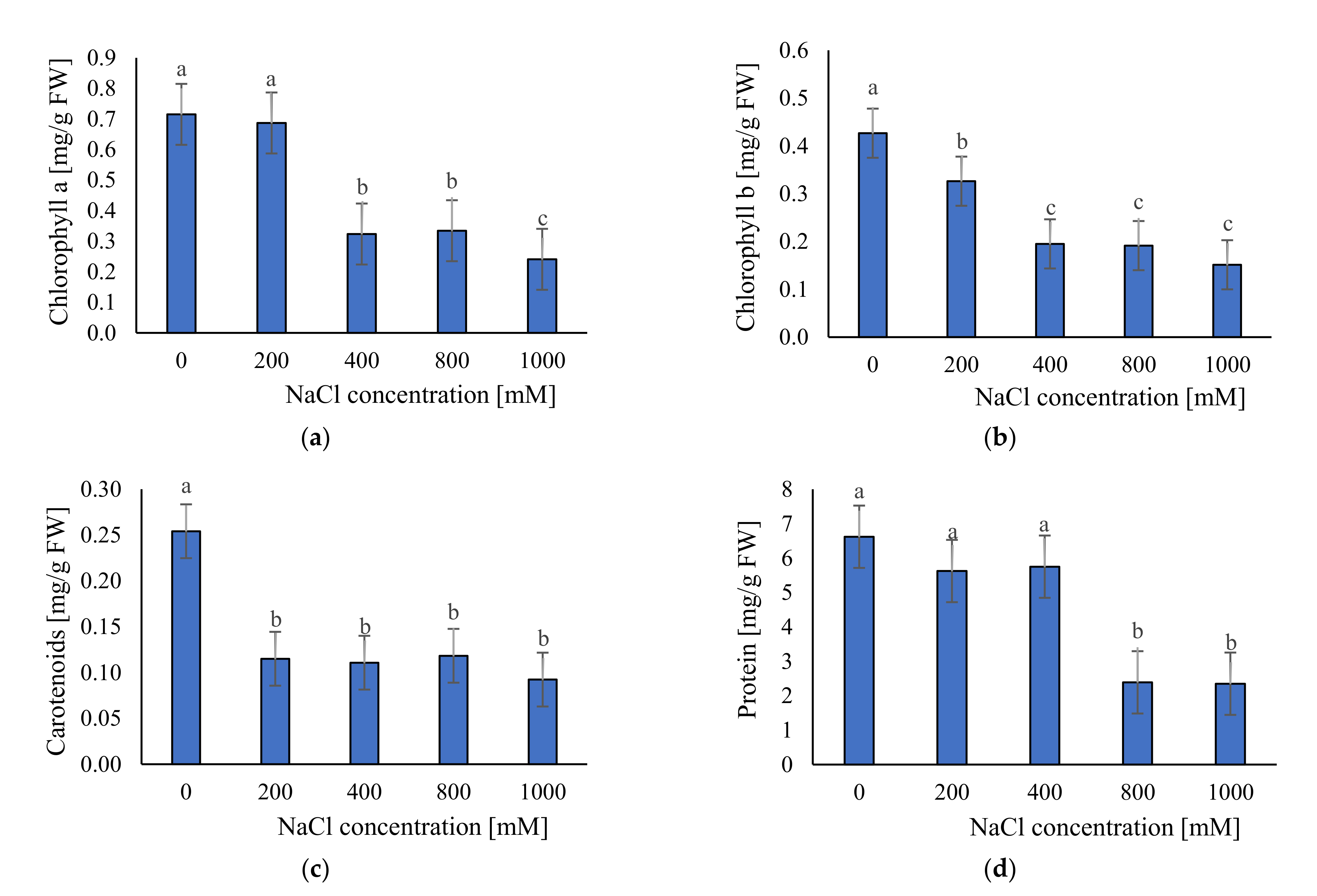
2.3.1. Photosynthetic Pigments and Soluble Protein Content
2.3.2. Hydrogen Peroxide (H2O2) Content
2.3.3. Antioxidant Enzymes Activities and Proline Content
3. Discussion
4. Materials and Methods
4.1. Laboratory Plant Material and Plantlets Preparation
4.2. Growth Analysis
4.3. Anatomical Analysis
4.4. Biochemical Analyzes
4.4.1. Photosynthetic Pigments Content
4.4.2. Soluble Protein Content
4.4.3. Hydrogen Peroxide (H2O2) Content
4.4.4. Peroxidase (POD) Activity
4.4.5. Catalase (CAT) Activity
4.4.6. Proline Content
4.5. Statistical Analyzes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a database of salt-tolerant plants: Helping put halophytes to work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscolo, A.; Panuccio, M.R.; Piernik, A. Ecology, Distribution and Ecophysiology of Salicornia Europaea L. In Sabkha Ecosystems; Khan, M.A., Böer, B., Öztürk, M., Al Abdessalaam, T.Z., Clüsener-Godt, M., Gul, B., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 47, pp. 233–240. [Google Scholar]
- Kadereit, G.; Ball, P.; Beer, S.; Mucina, L.; Sokoloff, D.; Teege, P.; Yaprak, A.E.; Freitag, H. A taxonomic nightmare comes true: Phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 2007, 56, 1143–1170. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas-Pérez, S.; Chanona-Pérez, J.J.; de Jesús Perea-Flores, M.; Calderon, H.; Piernik, A.; López-Soto, K.D.; González, C.B.G. Microstructure of Salicornia bigelovii stems under photonic and electron microscopy. Microsc. Microanal. 2020, 26, 360–362. [Google Scholar] [CrossRef]
- Cárdenas-Pérez, S.; Piernik, A.; Ludwiczak, A.; Duszyn, M.; Szmidt-Jaworska, A.; Chanona-Pérez, J. Image and fractal analysis as a tool for evaluating salinity growth response between two Salicornia europaea populations. BMC Plant Biol. 2020, 20, 467. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Loconsole, D.; Cristiano, G.; De Lucia, B. Glassworts: From wild salt marsh species to sustainable edible crops. Agriculture 2019, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Rezzouk, F.Z.; Thushar, S.; Shahid, M.; Elouafi, I.A.; Bort, J.; Serret, M.D. Effect of irrigation salinity and ecotype on the growth, physiological indicators and seed yield and quality of Salicornia europaea. Plant. Sci. 2021, 304, 110819. [Google Scholar] [CrossRef]
- Krüger, A.; Hellwig, F.; Oberprieler, C. Genetic diversity in natural and anthropogenic inland populations of salt-tolerant plants: Random amplified polymorphic DNA analyses of Aster tripolium L. (Compositae) and Salicornia ramosissima Woods (Chenopodiaceae). Mol. Ecol. 2002, 11, 1647–1655. [Google Scholar] [CrossRef]
- Antunes, M.D.; Gago, C.; Guerreiro, A.; Sousa, A.R.; Julião, M.; Miguel, M.G.; Faleiro, M.L.; Panagopoulos, T. Nutritional characterization and storage ability of Salicornia ramosissima and Sarcocornia perennis for fresh vegetable salads. Horticulturae 2021, 7, 6. [Google Scholar] [CrossRef]
- Rhee, M.H.; Park, H.-J.; Cho, J.Y. Salicornia herbacea: Botanical, chemical and pharmacological review of halophyte marsh plant. J. Med. Plants Res. 2009, 3, 548–555. [Google Scholar] [CrossRef]
- Cárdenas-Pérez, S.; Piernik, A.; Chanona-Pérez, J.J.; Grigore, M.N.; Perea-Flores, M.J. An overview of the emerging trends of the Salicornia L. genus as a sustainable crop. Environ. Exp. Bot. 2021, 191, 104606. [Google Scholar] [CrossRef]
- Liu, X.G.; Xia, Y.G.; Wang, F.; Sun, M.; Jin, Z.J.; Wang, G.T. Analysis of fatty acid composition of Salicornia europaea L. seed oil. Food Sci. 2005, 2, 42. [Google Scholar]
- Gunning, D. Cultivating Salicornia Europaea (Marsh Samphire); Irish Sea Fisheries Board: Dublin, Ireland, 2016; Volume 4, pp. 1–95. [Google Scholar]
- Patel, S. Salicornia: Evaluating the halophytic extremophile as a food and a pharmaceutical candidate. 3 Biotech 2016, 6, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigore, M.-N.; Toma, C. Morphological and anatomical adaptations of halophytes: A review. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Springer: Cham, Switzerland, 2021; pp. 1079–1221. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Hajibagher, M.A.; Yeo, A.R. Ion accumulation in the cell walls of rice plants growing under saline conditions: Evidence for the Oertli hypothesis. Plant Cell Environ. 1991, 14, 319–325. [Google Scholar] [CrossRef]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Effect of salt stress on physiological and antioxidative responses in two species of Salicornia (S. persica and S. europaea). Acta Physiol. Plant. 2011, 33, 1261–1270. [Google Scholar] [CrossRef]
- Mohammadi, H.; Kardan, J. Morphological and physiological responses of some halophytes to salinity stress. Ann. Univ. Mariae Curie Sklodowska Sect. C Biol. 2016, 70, 31. [Google Scholar] [CrossRef]
- Cárdenas-Pérez, S.; Niedojadło, K.; Mierek-Adamska, A.; Dąbrowska, G.B.; Piernik, A. Maternal salinity influences anatomical parameters, pectin content, biochemical and genetic modifications of two Salicornia europaea populations under salt stress. Sci. Rep. 2022, 12, 2968. [Google Scholar] [CrossRef]
- Mohamed, M.; El-Maboud, A. Seasonal Physiological Response of Salicornia Europaea L. Res. Rev. J. Bot. Sci. 2021, 10, 118–124. [Google Scholar]
- Akcin, A.; Yalcin, E. Effect of salinity stress on chlorophyll, carotenoid content, and proline in Salicornia prostrata Pall. and Suaeda prostrata Pall. subsp. prostrata (Amaranthaceae). Braz. J. Bot. 2016, 39, 101–106. [Google Scholar] [CrossRef]
- Rodríguez-Álvarez, M.; Ledea-Rodríguez, J.L.; Murillo-Amador, B.; Mazón-Suástegui, J.M. Morphometry and chlorophyll content of Salicornia bigelovii (Torr) under sea water treatments and Natrum muriaticum as a saline stress mitigator. Trop. Subtrop. Agroecosystems 2022, 25, 5. [Google Scholar]
- Wang, X.; Fan, P.; Song, H.; Chen, X.; Li, X.; Li, Y. Comparative proteomic analysis of differentially expressed proteins in shoots of Salicornia europaea under different salinity. J. Proteome Res. 2009, 8, 3331–3345. [Google Scholar] [CrossRef] [PubMed]
- Snow, A.A.; Vince, S.W. Plant zonation in Alaskan salt-marsh. II. An Experimental study of the role of edaphic conditions. J. Ecol. 1984, 72, 669–684. [Google Scholar] [CrossRef]
- Piernik, A. Growth of three meadow species along a salinity gradient in an inland saline habitat: Transplant experiment. Pol. J. Ecol. 2006, 54, 117–126. [Google Scholar]
- Lv, S.; Jiang, P.; Chen, X.; Fan, P.; Wang, X.; Li, Y. Multiple compartmentalization of sodium conferred salt tolerance in Salicornia europaea. Plant Physiol. Biochem. 2012, 51, 47–52. [Google Scholar] [CrossRef]
- Singh, D.; Buhmann, A.K.; Flowers, T.J.; Seal, C.E.; Papenbrock, J. Salicornia as a crop plant in temperate regions: Selection of genetically characterized ecotypes and optimization of their cultivation conditions. AoB Plants 2014, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.M.N.; Murty, P.P. Morphological and anatomical features of Salicornia brachiata Roxb. J. Biol. Chem. Res. 2013, 30, 887–891. [Google Scholar]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Enquist, B.J.; Condit, R.; Peet, R.K.; Schildhauer, M.; Thiers, B.M. Cyberinfrastructure for an integrated botanical information network to investigate the ecological impacts of global climate change on plant biodiversity. PeerJ Prepr. 2016, 4, e2615v2. [Google Scholar]
- Kattge, J.; Bönisch, G.; Dìaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zirbel, C.R.; Bassett, T.; Grmanand, E.; Brudvig, L.A. Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration. J. Appl. Ecol. 2017, 54, 1070–1079. [Google Scholar] [CrossRef] [Green Version]
- Bu, W.; Huang, J.; Xu, H.; Zang, R.; Ding, Y.; Li, Y.; Lin, M.; Wang, J.; Zhang, C. Plant Functional Traits Are the Mediators in Regulating Effects of Abiotic Site Conditions on Aboveground Carbon Stock-Evidence from a 30 ha Tropical Forest. Plot. Front. Plant Sci. 2019, 9, 1958. [Google Scholar] [CrossRef] [PubMed]
- Helsen, K.; Acharya, K.P.; Brunet, J.; Cousins, S.A.O.; Decocq, G.; Kolb, A.; Lemke, I.H.; Lenoir, J.; Plue, J.; Verheyen, K.; et al. Biotic and abiotic drivers of intraspecific trait variation within plant populations of three herbaceous plant species along a latitudinal gradient. BMC Ecol. 2017, 17, 38. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Cornelisson, J.H.C. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Raffard, A.; Lecerf, A.; Cote, J.; Buoro, M.; Lassus, R.; Cucherousset, J. The functional syndrome: Linking individual trait variability to ecosystem functioning. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171893. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Nie, L.; Jiang, P.; Feng, J.; Lv, S.; Chen, X.; Bao, H.; Guo, J.; Tai, F.; Wang, J.; et al. Transcriptome Analysis of Salicornia europaea under Saline Conditions Revealed the Adaptive Primary Metabolic Pathways as Early Events to Facilitate Salt Adaptation. PLoS ONE 2013, 8, e80595. [Google Scholar] [CrossRef]
- Kaleem, M.; Hameed, M. Functional traits for salinity tolerance in differently adapted populations of Fimbristylis complanata (Retz.). Int. J. Phytoremediation 2021, 23, 1319–1332. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Mann, A.; Lata, C.; Kumar, N.; Sharma, P. Salinity-induced physiological and molecular responses of halophytes. In Research Developments in Saline Agriculture; Springer: Singapore, 2019; pp. 331–356. [Google Scholar]
- Winicov, I.; Bastola, D.R. Salt tolerance in crop plants: New approaches through tissue culture and gene regulation. Acta Physiol. Plant. 1997, 19, 435–449. [Google Scholar] [CrossRef]
- Ulrich, W.; Kubota, Y.; Piernik, A.; Gotelli, N.J. Functional traits and environmental characteristics drive the degree of competitive intransitivity in European saltmarsh plant communities. J. Ecol. 2018, 106, 865–876. [Google Scholar] [CrossRef]
- Ulrich, W.; Hulisz, P.; Mantilla-Contreras, J.; Elvisto, T.; Piernik, A. Compensatory effects stabilize the functioning of Baltic brackish and salt marsh plant communities. Estuar. Coast. Shelf Sci. 2019, 231, 106480. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Salt stress effects on growth, pigments, proteins and lipid peroxidation in Salicornia persica and S. europaea. Biol. Plant. 2009, 53, 243–248. [Google Scholar] [CrossRef]
- Orlovsky, N.; Japakova, U.; Zhang, H.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers 2016, 38, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, H.; Caldeira, G.; Freitas, H. Salicornia ramosissima population dynamics and tolerance of salinity. Ecol. Res. 2007, 22, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Akcin, T.A.; Akcin, A.; Yalcın, E. Anatomical changes induced by salinity stress in Salicornia freitagii (Amaranthaceae). Braz. J. Bot. 2017, 40, 1013–1018. [Google Scholar] [CrossRef]
- Gong, D.H.; Wang, G.Z.; Si, W.T.; Zhou, Y.; Liu, Z.; Jia, J. Effects of salt stress on photosynthetic pigments and activity of ribulose-1, 5-bisphosphate carboxylase/oxygenase in Kalidium foliatum. Russ. J. Plant Physiol. 2018, 65, 98–103. [Google Scholar] [CrossRef]
- Rajabi Dehnavi, A.; Zahedi, M.; Ludwiczak, A.; Piernik, A. Foliar Application of Salicylic Acid Improves Salt Tolerance of Sorghum (Sorghum bicolor (L.) Moench). Plants 2022, 11, 368. [Google Scholar] [CrossRef]
- Ashraf, M. The effect of NaCl on water relations, chlorophyll, and protein and proline contents of two cultivars of blackgram (Vigna mungo L.). Plant Soil 1989, 119, 205–210. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Arulbalachandran, D.; Ganesh, K.S.; Subramani, A. Changes in metabolites and antioxidant enzyme activity of three Vigna species induced by NaCl stress. Am. Eurasian J. Agron. 2009, 2, 109–116. [Google Scholar]
- Ghanem, A.F.M.; Mohamed, E.; Kasem, A.M.M.A.; El-Ghamery, A.A. Differential Salt Tolerance Strategies in Three Halophytes from the Same Ecological Habitat: Augmentation of Antioxidant Enzymes and Compounds. Plants 2021, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B.; Mittra, B.; Mohanty, P. Salt-stress induced alterations in protein profile and protease activity in the mangrove Bruguiera parviflora. Z. Für Nat. C 2004, 59, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, A.C.; Duboué-Dijon, E.; Sterpone, F.; Hynes, J.T.; Laage, D. Biomolecular hydration dynamics: A jump model perspective. Chem. Soc. Rev. 2013, 42, 5672–5683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-T.; Qiu, Z.-B.; Zhang, X.-W.; Wang, L.-S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta Physiol. Plant. 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Liu, L.; Huang, L.; Lin, X.; Sun, C. Hydrogen peroxide alleviates salinity-induced damage through enhancing proline accumulation in wheat seedlings. Plant Cell Rep. 2020, 39, 567–575. [Google Scholar] [CrossRef]
- Hernandez, M.; Fernandez-Garcia, N.; Diaz-Vivancos, P.; Olmos, E. A different role for hydrogen peroxide and the antioxidative system under short and long salt stress in Brassica oleracea roots. J. Exp. Bot. 2010, 61, 521–535. [Google Scholar] [CrossRef] [Green Version]
- Shang, W.; Feierabend, J. Dependence of catalase photoinactivation in rye leaves on light intensity and quality and characterization of a chloroplast-mediated inactivation in red light. Photosynth. Res. 1999, 59, 201–213. [Google Scholar] [CrossRef]
- Amor, N.B.; Jiménez, A.; Megdiche, W.; Lundqvist, M.; Sevilla, F.; Abdelly, C. Response of antioxidant systems to NaCl stress in the halophyte Cakile maritima. Physiol. Plant. 2006, 126, 446–457. [Google Scholar] [CrossRef]
- Moghaieb, R.E.A.; Saneoka, H.; Fujita, K. Effect of salinity on osmotic adjustment, glycinebetaine accumulation and the betaine aldehyde dehydrogenase gene expression in two halophytic plants, Salicornia europaea and Suaeda maritima. Plant Sci. 2004, 166, 1345–1349. [Google Scholar] [CrossRef]
- Shoukat, E.; Ahmed, M.Z.; Abideen, Z.; Azeem, M.; Ibrahim, M.; Gul, B.; Khan, M.A. Short and long term salinity induced differences in growth and tissue specific ion regulation of Phragmites karka. Flora 2020, 263, 151550. [Google Scholar] [CrossRef]
- Pellegrini, E.; Forlani, G.; Boscutti, F.; Casolo, V. Evidence of non-structural carbohydrates-mediated response to flooding and salinity in Limonium narbonense and Salicornia fruticosa. Aquat. Bot. 2020, 166, 103265. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Rozema, J.; Schat, H. Salt tolerance of halophytes, research questions reviewed in the perspective of saline agriculture. Environ. Exp. Bot. 2013, 92, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Piernik, A. Ecological Pattern of Inland Salt Marsh Vegetation in Central Europe; Wydawnictwo Naukowe Uniwersytetu Mikołaja Kopernika: Toruń, Poland, 2012. [Google Scholar]
- Naidoo, G.; Naidoo, Y. Salt tolerance in Sporobolus virginicus: The importance of ion relations and salt secretion. Flora 1998, 193, 337–344. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Potential of Producing Salicornia bigelovii Hydroponically as a Vegetable at Moderate NaCl Salinity. HortScience Horts 2014, 49, 1154–1157. [Google Scholar] [CrossRef] [Green Version]
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Indicator values of plants in Central Europe. Scr. Geobot. 1992, 18, 1–248. [Google Scholar]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circular California Agricultural Experiment Station; University of California: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Cárdenas-Pérez, S.; Chanona-Pérez, J.; Méndez-Méndez, J.; Calderón-Domínguez, G.; López-Santiago, R.; Arzate-Vázquez, I. Nanoindentation study on apple tissue and isolated cells by atomic force microscopy, image and fractal analysis. Innov. Food Sci. Emerg. Technol. 2016, 34, 234–242. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. The assay of catalases and peroxidases. Methods Biochem. Anal. 1955, 1, 357–424. [Google Scholar] [CrossRef]
- Rached-Kanouni, M.; Alatou, D. Change in activity of antioxidative enzymes in leaves of Acacia retinodes, Biota orientalis and Casuarina equisetifolia under heat stress condition. Eur. Sci. J. 2013, 9, 402–410. [Google Scholar]
- Teranishi, Y.; Tanaka, A.; Osumi, M.; Fukui, S. Catalase activities of hydrocarbon-utilizing Candida yeasts. Agric. Biol. Chem. 1974, 38, 1213–1220. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditional Effects | |||
|---|---|---|---|
| Variable | V % | Pseudo-F | p |
| Chl a | 24.8 | 7.2 | 0.002 |
| Carot | 24.7 | 10.3 | 0.002 |
| Psurf | 24.5 | 18.8 | 0.002 |
| Protein | 23.1 | 147 | 0.002 |
| POD | 0.8 | 6.5 | 0.014 |
| Nbran | 0.6 | 6.4 | 0.012 |
| AR | 0.7 | 11.3 | 0.004 |
| Cperim | 0.2 | 3.9 | 0.03 |
| Proline | <0.1 | 1 | 0.372 |
| CAT | <0.1 | 1.4 | 0.286 |
| H2O2 | <0.1 | 0.8 | 0.516 |
| Cdiam | <0.1 | 1.3 | 0.304 |
| Sdiam | <0.1 | 0.6 | 0.598 |
| Carea | <0.1 | 1 | 0.396 |
| SFW | <0.1 | 0.9 | 0.452 |
| TDW | <0.1 | 1.2 | 0.334 |
| Pheight | <0.1 | 0.6 | 0.624 |
| Chl b | <0.1 | 2.2 | 0.144 |
| RFW | <0.1 | 0.7 | 0.5 |
| SDW | <0.1 | 0.8 | 0.466 |
| Cround | <0.1 | 0.2 | 0.852 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cárdenas-Pérez, S.; Rajabi Dehnavi, A.; Leszczyński, K.; Lubińska-Mielińska, S.; Ludwiczak, A.; Piernik, A. Salicornia europaea L. Functional Traits Indicate Its Optimum Growth. Plants 2022, 11, 1051. https://doi.org/10.3390/plants11081051
Cárdenas-Pérez S, Rajabi Dehnavi A, Leszczyński K, Lubińska-Mielińska S, Ludwiczak A, Piernik A. Salicornia europaea L. Functional Traits Indicate Its Optimum Growth. Plants. 2022; 11(8):1051. https://doi.org/10.3390/plants11081051
Chicago/Turabian StyleCárdenas-Pérez, Stefany, Ahmad Rajabi Dehnavi, Karol Leszczyński, Sandra Lubińska-Mielińska, Agnieszka Ludwiczak, and Agnieszka Piernik. 2022. "Salicornia europaea L. Functional Traits Indicate Its Optimum Growth" Plants 11, no. 8: 1051. https://doi.org/10.3390/plants11081051